@kanri_ninjin
@kanri_ninjin


 イクナイ! 581
イクナイ! 581







2 活動電位
前節では神経細胞の形態と、 その基本的な性質である静止膜電位について解説した。 とくにイオン平衡による静止膜電位の発生のメカニズムは、 ニューロンの電気的応答を理解するための基礎であるため、 しっかりと理解してほしい。
といっても、 じつは「膜電位をもつ」という特徴は、 ほとんどの生体細胞にみられる一般的な性質である。 どんな細胞でも内部と外部には少なからずイオン濃度差があり、 多少ともイオンを透過する細胞膜でおおわれているからだ。 ニューロンの真骨頂は、 その電位を一過性に変化させることができる点にある。 これを活動電位というのだった。 本節では、活動電位の詳しい発生機序をみていこう。
2.1 電位依存性Na+チャネル
ここまで、 静止膜電位がK+の濃度勾配と選択的透過性により生じ、 その値は膜電位に対するK+の寄与が他のイオンよりも優位になることで、 K+平衡電位に近くなっていることを学習した。 ではたとえば、 ニューロンの細胞膜に「扉」のようなものがあって、 それが突然開くことによって他のイオンの平衡電位の寄与が一気に増大したらどうなるだろうか。
たとえばNa+を考えよう。 もしNa+の膜透過性が一気に増大すると、 Na+の平衡電位が膜電位にあたえる寄与が大きくなり、 細胞内の電位はNa+の平衡電位(+59mV)に近づくだろう。 じつはこれこそが活動電位発生のメカニズムそのものである。 本項ではこの過程の分子的な実態をひも解いていこう。
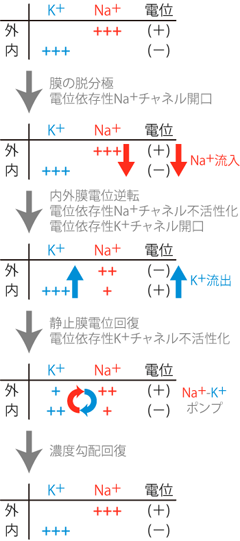
まず上で説明した「扉」とは何なのか、 それが分からないことには話が進まない。 膜上に存在してイオンを通すのは、 これまでもみてきたとおりチャネルタンパクの十八番である。 実際、活動電位の発生にも、 イオンチャネルが重要な役割を果たす。 ただしそのチャネルは、 K+リークチャネルのようにいつでも開いたままではダメだ。 活動電位は、ニューロンが他のニューロンに情報を伝達する際に発する、 パルス的な電気信号なのだ。 よってその活動電位を発生するためのチャネルは、 情報を伝達する際にのみ開き、他の場合には閉じていなければならない。 そこでニューロンは 電位変化を検知して開閉する 電位依存性Na+チャネル voltage-dependent sodium channelを用い、 活動電位の発生を制御している。 電位依存性Na+チャネルの模式図をFigure 7に示す。
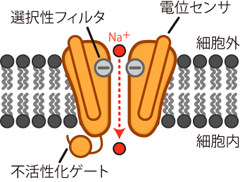
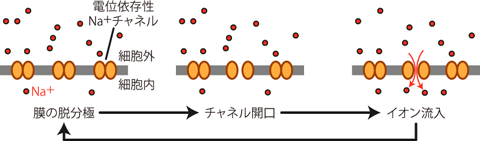
このチャネルは、 普段は閉じていてイオンを通さないが、 ある一定以上の正の電位変化(すなわち脱分極)によって立体構造の変化を起こし、 Na+が中心の穴を通れるようになる。 その結果、 静止状態では膜に阻まれて細胞内に入ってこられないNa+が、 電気的勾配および化学的勾配の双方の力によって細胞内に流入する。 (Na+は細胞内より細胞外に多く、 また電気的にもマイナスの細胞内に引きつけられる。) こうして陽イオンが細胞内に入ってくる結果、細胞は少し脱分極する。
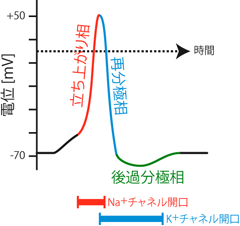
この脱分極性(すなわちプラス方向)の膜電位変化は、 膜上のまだ開いていなかった電位依存性Na+チャネルをさらに開口させる。 そしてそのチャネルから細胞内へとさらにNa+が流入し、 細胞はより脱分極する。 このような正のフィードバック機構により、 細胞膜上の微小部分に存在する電位依存性Na+チャネルが一気に開口することになる。 すなわちその部分では膜電位に対するNa+平衡電位の寄与が一時的に増大するため、 膜電位がNa+の平衡電位に近い+50mV程度まで一気に上昇し、 膜内外電位の正負が瞬間的に逆転するのである。 このような細胞膜の急激な電位変化を興奮 excitationという。 電位依存性Na+チャネルの正のフィードバックによる急激な膜電位上昇機構を、 発見者の名前にちなんでホジキンサイクル Hodgkin cycleとよぶ (Figure 8)。
このホジキンサイクルによって引き起こされる膜興奮こそが、 活動電位発生の第一段階である。
2.2 全か無かの法則
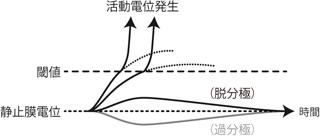
先ほどこの機構の要である電位依存性Na+チャネルが 「ある一定以上の正の膜電位変化を感知して開く」と書いた。 逆にいえばいくら膜が脱分極しても、 それがこのチャネルの開口を誘発するだけの電位変化 (「ある一定以上」の膜電位変化)でなければ、 ホジキンサイクルによる急激な電位上昇は起こらない。 このような電位上昇を引き起こしうる脱分極の「ボーダーライン」のことを 閾値 thresholdという。
この機構により、閾値に達した脱分極はそれがどの程度の大きさのものだろうと、 必ずホジキンサイクルによる膜内外の電位逆転を引き起こす。 逆も同様であり、 閾値に達しなかった脱分極はやはりその程度に関わらず結局静止膜電位に戻ってしまい、 何もなかったのと同じことになる。
このように細胞膜のある部分に与えられた小さな膜電位変化は、 それが閾値に「達するか/達しないか」によって、 膜電位の急上昇を「起こすか/起こさないか」のどちらかの結果になる。 閾値の半分程度の電位変化が起こったからといって、 その結果Na+平衡電位の半分程度の大きさまで、 ほどほどな電位上昇を生じるということはない。 これを全か無かの法則 all-or-none theoryといい、 細胞によるこのような反応を 全か無かの反応 all-or-none responseと呼ぶ。 すなわち活動電位はその分子的な発生機序の特性により、 発生するときは一定に発生するし、 発生しないときはまったく変化がないというon/off変化をすることになる (Figure 9)。
2.3 電位依存性K+チャネル
電位依存性Na+チャネルのはたらきで膜電位が急上昇することにより、 活動電位発生の最初のプロセスは達成された。 しかし活動電位は、 ニューロンにおける細胞内電位の「一過的」変化のはずだった。 もしこのままNa+が細胞内に流れ込み続けては、 ニューロンはいつまでたっても静止膜電位に戻れず、 電位変化が「一過性」にはならない。
そこで急激な細胞内電位の上昇をストップさせるため、 電位依存性Na+チャネルには不活性化という機能が備わっている。 電位依存性Na+チャネルの不活性化とは、 ホジキンサイクルによって一気に活性化(開口)した電位依存性Na+チャネルが、 開口から1ミリ秒もしないうちに更なる立体構造変化によって口を閉じ、 Na+を通さなくなることである。 こうしてNa+の細胞内への急激な流入は止まる。
しかしこれだけでは、まだ急激なNa+の流入が止まっただけで、 細胞内はプラスのままだ。 よってこの変化を「一過性」にするためには、 細胞内電位を再び静止膜電位レベルにまで下げなおさなければならない。 このプロセスを細胞の再分極 repolarizationと呼ぶ。 神経細胞においては、 電位依存性K+チャネル voltage-dependent potassium channel が再分極の役割を担っている。
電位依存性Na+チャネルにより細胞内がプラスになったとき、 K+にはたらく駆動力(K+を移動させようとする力)を考えてみよう。 まずK+の濃度だが、 これは静止膜電位のときと変わらず細胞内のほうが外より非常に大きい。 一方、電気的な状態はというと、 (何度もくどくて申し訳ないが)一時的に細胞内が正で細胞外が負になっている。 よって細胞内のK+には、電気的にマイナスな細胞外へ引っ張る力と、 濃度勾配の力によって外へと押し出す力の両方がかかることとなる。
電位依存性K+チャネルは、 まさにこのタイミングで、 Na+チャネルの不活性化に合わせるようにして一斉に開口する。 これによって普段以上のK+透過性が膜に生じると、 K+は一気に細胞外へと排出され、 結果的に細胞内は陽イオンを失ってマイナスに変化する。 これが逆転した細胞内外の電位差を元の状態にまで戻す再分極の機構である。 あとは先述のNa+-K+ポンプが、 ATPのエネルギーを用いてNa+とK+の濃度勾配を回復すれば、 これでめでたく膜内外の環境はもとの静止状態に戻る。 ここまでの機構をFigure 10に示す。
ちなみに電位依存性K+チャネルというくらいだから、 このチャネルも電位依存性Na+チャネルと同じく正の電位変化を受容して開口する。 しかしこのチャネルがNa+チャネルと同時に開いたら、 Na+流入とK+流出が相殺して活動電位が発生しなくなってしまうかもしれない。 そこで 電位依存性K+チャネルは、 電位依存性Na+チャネルの開口に少し遅れて開くようになっている。 また逆に電位依存性K+チャネルは、 Na+チャネルが閉じたあとも膜の電位をマイナスに戻すために働かなければならないから、 電位依存性Na+チャネルが不活性化したあともしばらく働き続ける (Figure 11)。 このような電位依存性イオンチャネルの開閉のタイムラグにより、 急激な立ち上がりと続く再分極による鋭い電位変化が生じる。 ニューロンの活動電位はこのような波形から スパイク spikeと表現されることもある。 またその一過的な性質からインパルス impulseとも呼ばれる。 もともと分極状態であった細胞が瞬間的に脱分極することから、 ディスチャージ discharge(「放電」の意)とも表現する。 これらはいずれも「活動電位」の同義語である。