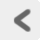@kanri_ninjin
@kanri_ninjin


 イクナイ! 581
イクナイ! 581







3 脳
前節では、脊髄(中枢神経系) および脊髄から出る脊髄神経(末梢神経系)の解剖についてみてきた。 ここからは中枢神経系を構成するもうひとつの要素である脳と、 そこから出る末梢神経である脳神経について学習していこう。
3.1 脳の分類
脳は大脳 cerebrum・ 間脳 interbrain・ 中脳 midbrain・ 橋 pons・ 延髄 medulla oblongata・ 小脳 cerebellumで構成されている(Figure 15a)。 これらのうち中脳・橋・延髄を合わせて 脳幹 brainstemと呼ぶこともある。 これはこの部分が、脳全体を木に見立てたとき、 その幹を構成する部分だからである(Figure 15b)。 脳幹という呼称は便利なため多用されているが、 間脳を含む場合と含まない場合があるので、 それが指している範囲は文脈によって柔軟に理解する必要がある。
3.2 脳の支持構造
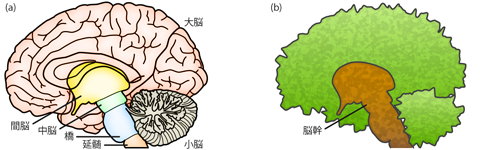
ヒトの頭部は(耳小骨をのぞくと)22個の骨でできている。 これらの骨の強固な結合によってつくられる骨格構造、 いわゆる頭蓋骨(ずがいこつ)を頭蓋 cranium(とうがい)という。 (前述のとおり、解剖学用語は音読みする慣習となっている。) 頭蓋を構成する22の頭蓋骨(とうがいこつ)のうち、 14個は顔面や顎にあたる顔面頭蓋 facial craniumをつくる。 残る8つ(後頭骨・左右頭頂骨・前頭骨・左右側頭骨・蝶形骨・篩骨)は、 ドーム状に組み合わさって脳を取り囲む 脳頭蓋 cranial bonesをつくる(Figure 16)。 脳頭蓋内部の中空の部分を頭蓋腔 cranial cavityといい、 脳はここにおさめられている。
とはいえもちろん、 頭蓋腔に直に脳が入っているわけではない。 脊髄が脊髄髄膜で覆われていたのと同様、脳も3枚の 脳髄膜 cranial meninx(硬膜・クモ膜・軟膜)で 覆われている。 脊髄においては脊柱管と硬膜のあいだに詰め物となる脂肪組織等が充填されていたが、 脳においては、 硬膜は直接脳頭蓋の骨内壁に癒着している。 これは姿勢によっておおきく形態が変わる脊柱管と違い、 脳頭蓋の形態はほぼ変化しないため、 「あそび」をつくらずに付着しても内部の神経組織を痛める危険がないからである。 硬膜の内層にはクモ膜があり、 両者のあいだにはすきまはない。 (死後解剖では硬膜とクモ膜のあいだに硬膜下腔がみとめられるが、 この空間は生体にはないと考えられている。) クモ膜とさらに内層の軟膜のあいだには、 脊髄同様クモ膜下腔が存在し、 脳脊髄液で満たされている。
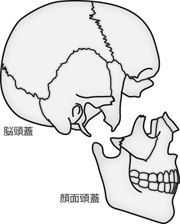
3枚の髄膜のうちもっとも内層の軟膜は、 脳にべったりと張り付き、 脳表面のミゾの内部まで入り込んでいる。 一方で硬膜とクモ膜は、 基本的に脳表面にあるミゾは無視するかたちで、 脳全体を大まかに包んでいる。 しかし左右の大脳半球のあいだや、大脳と小脳のあいだなどの広いすきまには、 軟膜だけでなく硬膜とクモ膜が入り込み、 脳全体を内部から支える仕切り板の役割を果たしている。 左右の大脳半球のあいだを仕切る膜構造を 大脳鎌 falx cerebriという(Figure 17)。 これは左右半球間に入り込んだ硬膜の三日月状の形状が、 鎌のように見えることに由来する。 同様に左右の小脳半球のあいだを仕切る膜構造を 小脳鎌 falx cerebelliという。 小脳は半球間のミゾが浅いため、 小脳鎌は大脳鎌と比べるとかなり細い。 さらに、大脳と小脳のあいだを仕切る膜を 小脳テント tentorium cerebelliと呼ぶ。 これはこの膜が、 ちょうど小脳の頂点を天蓋として、 小脳背面を覆うかたちに吊ったテントのようにみえることによる。
大脳鎌・小脳鎌と小脳テントの3次元形態はイメージしにくいので、 もう少し説明を加えよう。 頭部を正中線からほんの少し左右どちらかにずらして矢状断面にしたとする。 すると、 アジの開きの片身側に背骨が残るように、 左右の半球間にはいりこんだ正中の硬膜・クモ膜は片側にだけ残り、 もう半面には脳の内側面が露出することになる(Figure 17)。 このとき観察できる三日月状の部分が大脳鎌と小脳鎌だ。 またここからさらに小脳をはずしてみると、 小脳の上面と大脳の下面に挟まれていたエンペラ状の膜が確認できる。 この部分が小脳テントである。 先の魚の例でいうと、 小脳を魚の「ワタ」の部分とみなしたとき、 身とワタとを区切っている腹骨にあたるのが小脳テントだ。 このような大きなミゾに入り込んだ硬膜とクモ膜は、 脳を内側から支持して安定させる役割を果たしている。
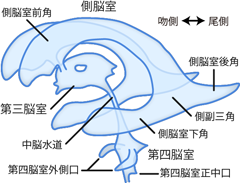
さて、今度は脳の内部構造へと目を向けよう。 横断面でみた際、脊髄の中芯部分には中心管と呼ばれる空間が存在した。 このような神経組織内部の空洞は脳にも存在し、 脳室 ventricleと呼ばれている。 脳室の空洞に型材を流し込んでキャストを取ったとすれば、 Figure 18のような複雑な構造が観察できる。 ただし脳室だけ眺めていても周囲の神経構造との位置関係が分かりづらいので、 後述するFigure 20のイラストも並べてみながら確認してほしい。 ヒトの脳には、 左右大脳半球のなかの側脳室 lateral ventricle、 間脳のあいだの第三脳室 third ventricle、 橋のうしろの第四脳室 fourth ventricleという、 計4つ(側脳室2つ+第三脳室+第四脳室)の脳室がある。 番号付きの脳室がいきなり「第三」からはじまるのは、 左右の側脳室を1・2とみなすためである。 脳室の内部は、脊髄の中心管と同様、脳脊髄液で満たされている。 側脳室と第三脳室は室間孔 interventricular foramen (モンロー孔 foramen of Monro)でつながっているが、 側脳室同士は直接つながってはいない。 すなわち正中に位置する第三脳室から、 左右の側脳室へとのびた渡り廊下が室間孔だ。 さらに第三脳室は、 後端部分の中脳水道 mesencephalic aqueduct により第四脳室とつながっている。 中脳水道は、名前の通り中脳のレベルにあたる。 さらに第四脳室は、 その後端においてそのまま脊髄レベルに到り、 脊髄の中心管へと移行している。
3.3 脳脊髄液系
本節では、脳の内外を満たす脳脊髄液の動態について説明しよう。
先に述べたとおり、脳と脊髄にはどちらも内部に空洞の部分が存在する。 脳内にある空洞を脳室、脊髄内にある空洞を中心管といった。 これらの空洞は別々のものではなく、 左右側脳室と第三脳室が、 第三脳室と第四脳室が、 そして第四脳室と中心管がつながるかたちで、 全体でひと続きになっていた。 そしてその内部は、脳脊髄液という液体で満たされているのだった。
一方、中枢神経系を包む髄膜のうちクモ膜と軟膜のあいだには、 クモ膜下腔と呼ばれる空間が存在した。 このクモ膜下腔は脳にも脊髄にも存在し、 やはり脳のクモ膜下腔と脊髄のそれはひと続きになっている。 この中枢神経系を取り囲んだクモ膜下腔も脳脊髄液で満たされており、 これにより脳と脊髄は、 クモ膜で包まれた脳脊髄液のプールのなかに浮かんだ状態になっている。 この液体のクッションにより、 脳は外からかかる衝撃から守られている。 また液体内に浮いているために、 脳の見かけの重量は50g程度におさえられ、 自重でつぶれるのを免れている。 (実際の脳の重さは1400gほどある。)
ところで脳脊髄液には、栄養分や老廃物など、 さまざまな化学物質が溶け込んでいる。 すなわち脳脊髄液はクッションの働きだけでなく、 中枢神経系の内外から神経組織を栄養したり、 老廃物を除去したりする役割をもっている。 よって脳脊髄液は単に空洞を満たすだけではなく、 つねに代謝・循環され、 新鮮な状態に保たれなければならない。
脳脊髄液は、 脳室にある脈絡叢 choroid plexus という組織で産生される。脈絡叢とは、脳室表面の細胞と毛細血管が複合した微細なヒダ状の組織で、 ここから浸み出した血漿成分が脳脊髄液である。 当然ながら赤血球などの細胞は毛細血管壁をすり抜けることはできない。 また脈絡叢では血管をおおう脳室表面の細胞が特殊化し、 受動輸送・能動輸送を駆使して、 血液から脳脊髄液に浸み出す成分を制限している。 たとえば血中に存在するアミノ酸であるグリシンは、 神経細胞に対して神経伝達物質としてはたらいてしまう。 そのためグリシンは脳脊髄液へと浸み出すことを許されず、 脳脊髄液中では原料となる血中よりも非常に低い濃度に保たれている。
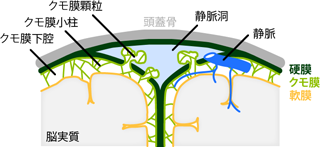
脈絡叢は第三脳室の天井部分に貼りつくように存在し、 室間孔を通って側脳室の底面にも広がっている。 また第四脳室の背部にも存在する。 脳脊髄液はこれらの場所でつねに血液から浸み出し続けていて、 一日あたり約500mlが産生される。 しかしつくられるばかりでは、 すぐに脳室・中心管がパンクしてしまう。 そうならないためには、 古くなった脳脊髄液を、 産生と同じペースで除去しなければならない。 その役を担うのが硬膜静脈洞 dural sinusである (Figure 19)。 硬膜静脈洞では、脳をつつむ硬膜が部分的に内層と外層の2枚に分かれ、 その間に静脈が入り込んでいる。 また静脈洞のクモ膜下腔側の表面では、 クモ膜が部分的に硬膜を貫いて静脈洞内へと陥入している (クモ膜顆粒 arachnoid granulation)。 もともとが血漿成分である脳脊髄液は、 クモ膜顆粒において静脈へと回収され、 血流によって代謝される。
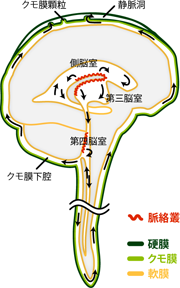
それでは脳脊髄液の循環の様子を、 順を追って確認してみよう(Figure 20)。 脳脊髄液は、脳室の脈絡叢で産生される。 そしてゆっくりと脳室内を第四脳室に向かって流れていく。 ごく少量の脳脊髄液は、脊髄の中心管内にも流れ込み、 ふたたび第四脳室に向かってもどってくる。 第四脳室には正中孔(マジャンディー孔)と外側孔(ルシュカ孔)という穴が存在する。 これは脳室とクモ膜下腔をつなぐ唯一のトンネルである。 この穴を通って、脳脊髄液は脳室からクモ膜下腔へと流れ出る。 脳脊髄液はさらにクモ膜下腔をゆっくりと循環していき、 最終的に頭蓋内の静脈洞へと再吸収される。
このように脳室とクモ膜下腔を満たす脳脊髄液の実態は、 パイプを介して連絡するプールに満たされた同じ液体である。 脳脊髄液は脳室の脈絡叢で産生され、 脳室・中心管とクモ膜下腔を循環したのち、 硬膜内を走る静脈へと吸収されていくのである。 この産生と回収が一日500mlずつ行なわれているおかげで、 われわれの中枢神経系は、 いつも新鮮な脳脊髄液のなかに全身浴していられるわけだ。
3.4 脳の動脈
前項では、 中枢神経系を物理的に保護すると同時に、 神経組織の栄養と代謝にも関わる脳脊髄液循環について紹介した。 本項では、 より直接的に神経系へと酸素や栄養を届ける血液循環のシステムについて学習しよう。
脳はとても「ぜいたく」な器官である。 ヒトの脳が全体重に占める割合はたった2%にすぎないが、 脳が使うエネルギーや酸素の量は個体全体の約20%にも達する。 カロリーにすると約500kcalものエネルギーを、 脳は毎日消費しているのだ。 一方で脳は大喰らいなだけでなくグルメでもある。 というのも神経組織は、 主たる栄養源としてグルコースのみを使い、 脂肪酸は利用しない。 そのくせ脳自身はみずからグルコースの貯蔵が全くできないので、 つねにグルコースの補給を受け続けなければならない。 もちろんそのグルコースを利用するためには、 当然酸素の供給も必要である。
このように脳が正常に働くためには、 栄養と酸素を供給するための血液循環が不可欠である。 たった10秒でも脳への血液供給が途切れると、 脳は正常な機能ができなくなり、 われわれの意識は遠のいてしまう。 また脳内のほんの一部の血管に病変が生じただけでも、 その血管が栄養していた脳領域が機能できなくなり、 部位によってさまざまな障害が生じる。
新鮮な血液から栄養と酸素を受けとるため、 脳の表面は血管によっておおわれている。 とくにヒトの脳の場合、 大きく発達した大脳皮質全体をもれなくカバーする必要がある。 この大脳の血液循環には、 前大脳動脈 anterior cerebral artery・ 中大脳動脈 middle cerebral artery・ 後大脳動脈 posterior cerebral artery という3種の動脈が関わっている。 これらの動脈は大脳を栄養する3本(左右で6本) の主要幹線道路にあたる。 このメインストリートから大小さまざまな分枝がわかれ、 毛細血管となって脳組織へと栄養を届けるわけだ。 しかしこれら3種の動脈の走行経路をみるに先立ち、 まずは頚部からこれらの大脳動脈へと到るまでの血液の供給ルートをおさえておこう。
いうまでもなく血液は心臓から送り出されて各器官へと届けられる。 脳へと向かう動脈は、 心臓を出た直後に大動脈から分岐して上方へ向かう。 上半身の広範な領域へと向かうこれらの分枝のうち、 頭部へ血液を供給する血管は 椎骨動脈 vertebral arteryと 総頚動脈 common carotid arteryの2種類である。 椎骨動脈は頚椎の横を、 総頚動脈は気管の横を上行するので、 総頚動脈のほうがやや腹側にある(Figure 21)。 頚部の椎骨には椎体の左右に横突孔という小さな穴が開いており、 椎骨動脈はこれを通って、 頚椎をビーズどめするようなかたちで頭部へと向かう。 一方で総頚動脈はそのように骨に守られてはいないので、 気道のわきをおさえるように指をあてるとその脈動を感じることができる。 総頚動脈は頭部に入る前に、 さらに内頚動脈 internal carotid arteryと 外頚動脈 external carotid arteryに分かれる。 このうち外頚動脈は顔面や頭部の皮膚へ向かうため、 脳の栄養には関与していない。 残る内頚動脈が椎骨動脈とともに頭蓋内に入り、 脳へと血液を届ける。
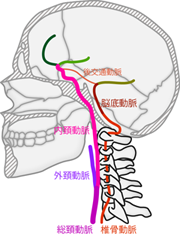
椎骨動脈と内頚動脈では頭蓋腔に入る際のルートも異なる。 脊椎に沿って上行した椎骨動脈は、 脊柱管内の脊髄と同じく、 頭蓋骨と脊椎の接合部の 大後頭孔 foramen magnumから頭蓋腔に入る。 頭蓋腔に入った左右の椎骨動脈は延髄のレベルで合流し、 正中を走る1本の脳底動脈 basilar arteryとなる。 脳底動脈は橋の腹側に貼りつくようにしてさらに上行する。 一方で内頚動脈は、 左右の骨にあいた頚動脈管 carotid canal という専用の穴を通って頭蓋腔に入る。 頚動脈管は側頭骨のなかを前内側へと漸進しながら上へと続いている。 内頚動脈はそのなかを通り、 視床下部の脳下垂体と呼ばれる構造の両脇あたりで頭蓋腔内へと顔をだすことになる。
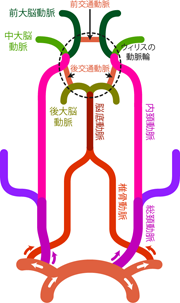
これでやっと頭蓋腔内に到達した。 Figure 22のダイアグラムをみながらここまでの流れを確認し、 整理ができたら先へと進もう。 内頸動脈は頭蓋内に入ると、 大脳を栄養する前述の3種の動脈(前・中・後大脳動脈)のうち、 前大脳動脈と中大脳動脈とに分岐する。 一方で(左右椎骨動脈が合流した)脳底動脈は、 中脳の腹側あたりでふたたび左右にわかれて後大脳動脈になる。 このように大脳をカバーする前・中・後3本の幹線道路のうち、 前2者は内頚動脈に、 後者は脳底動脈に由来する。 しかし同時に、 同側の中大脳動脈と後大脳動脈は 後交通動脈 posterior communicating artery によってつながっている。 さらに左右の前大脳動脈も、 前交通動脈 anterior communicating artery によって連絡されている。 言い換えれば左右6本の前・中・後大脳動脈は、 その基部において輪をつくるようにしてつながっている。 これを大脳動脈輪 cerebral arterial circle (ウィリスの動脈輪 circle of Willis)という (Figure 22)。 このようなバイパス構造の存在により、 動脈の一部に不具合が生じて血流が妨げられたとしても、 他のルートから回り込んで血液が届けられ、 深刻な影響が生じにくいようになっている。
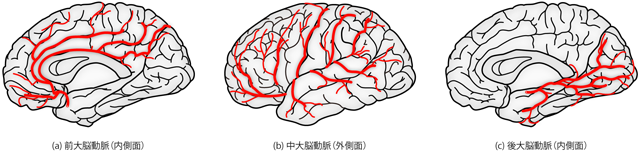
こうしてそれぞれのルートから大脳へと達した前・中・後大脳動脈は、 各々おおよそ決まった範囲の皮質へと枝を伸ばし、 その範囲を栄養する(Figure 23)。 前大脳動脈と後大脳動脈はおもに皮質内側面の吻側・尾側領域をそれぞれ担当する。 これらの内側面の動脈の一部は、 大脳のふちをまわりこんで外側面の外縁部の栄養も行なう。 一方で中大脳動脈は、 前頭葉と側頭葉のあいだの外側溝と呼ばれる隙間から回り込み、 外側面の広い領域を栄養する。 また脳脊髄液循環の解説において、 脳脊髄液は脈絡叢という組織で血液から産生されることを学習した。 脳脊髄液は新鮮な血液からつくらなければならないから、 脈絡叢は当然、 動脈から酸素と栄養素に富む動脈血の供給を受ける必要がある。 こうした脈絡叢や脳幹・小脳などの栄養は、 脳底動脈が上行する過程で別れ出た数多くの動脈の分枝が担っている。 脳部位の名称についてはまだ解説していないので、 後続の項において学習が済んでから、 改めて本項に立ち戻り、 各動脈の担当領域の違いを確認してほしい。
3.5 脳の静脈
前項の動脈に引き続き、 本項では静脈についても簡単にみておこう(Figure 24)。 頭蓋内に入った動脈は分枝しながらどんどん細くなっていき、 毛細血管となる。 毛細血管は養分や酸素を神経組織に与えたあと、 ふたたび集まって静脈を形成する。 静脈は動脈とおなじく、 頭蓋腔内で脳の表面を走っている。 こうした静脈はさらに合流し、 硬膜静脈洞へと集合する。
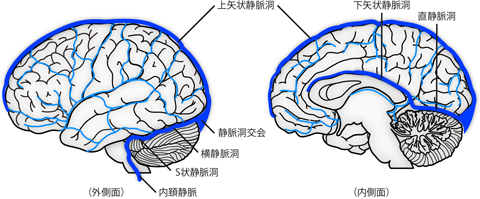
脳脊髄液系の項で説明したとおり、 静脈洞は、 内層と外層にわかれた硬膜のあいだに静脈がつつまれたような構造だった (Figure 19)。 柔軟に敷きまわせる普通の静脈と違い、 静脈洞は特殊化した硬膜をともなう構造なので、 その配置はかなり固定的で個人差が少ない。 大脳鎌の上端、 すなわち頭蓋冠(あたまのてっぺん)の硬膜とのつけ根部分を前後に走る 上矢状静脈洞 superior sagittal sinusは、 最大の硬膜静脈洞である。 上矢状静脈洞内では、 血液は前方から後方へと流れている。 また大脳鎌の下端にも、 脳梁の上を這う位置に 下矢状静脈洞 inferior sagittal sinus が通っている。 下矢状静脈洞はその後端で小脳テントの「稜線」を通る 直静脈洞 straight sinusへと続いており、 やはり前方から後方へと静脈血を集めてくる。 これらの正中の静脈洞は後頭骨のでっぱり(外後頭隆起)の直下あたりで合流し(静脈洞交会)、 左右の横静脈洞 transverse sinusへとわかれる。 横静脈洞は大脳と小脳のあいだの外側面、 すなわち小脳テントの「裾」をたどるように前側方へと回り込む。 そして小脳の前端あたりで脳幹部に寄り添うように内側へ引き寄せられ (S状静脈洞 sigmoid sinus)、 内頚静脈となって頭蓋腔を出ていく。
このように静脈洞から集められた静脈血は、 最終的に内頚静脈へと集まる。 内頚「動」脈が頚動脈管という骨にあいた専用の穴を通ったのと同様に、 内頚「静」脈も頚静脈孔という専用トンネルを通って頚部へと抜ける。 脳からの血液の流出のメインルートは、 この左右計2本の内頚静脈のみである。 内頚動脈と椎骨動脈という2人の役者のいた動脈とのアナロジーから 椎骨静脈 vertebral vein の関与も期待してしまうが、 椎骨静脈はおもに頭蓋外からの静脈血を運んでおり、 脳からの血液流出にはほとんど関わっていない。
前項と本項では、 脳内における血液循環の経路を、 おもに大脳皮質の栄養に関与する大きな動脈・静脈にしぼって学習した。 このような脳内を走る血管の分布や名称は、 神経系の解剖学そのものと比べると脇役的なトピックである。 そのため入門者向けの解説書などでは、 こうした神経系の代謝に関する基礎知識の優先度は低く、 ほとんど扱っていない教科書も多い。 しかし脳梗塞や脳内出血など、 血管性の脳機能障害について考えるとき、 脳に張り巡らされた血管の分布や名称の知識は不可欠である。 とくに脳へ血液を供給する動脈に関しては、 その損傷が下流の脳部位の虚血に直結しているため、 どの血管がどの領域の栄養を担っているか即座にわかるようにしておくことは重要である。 だから本解説文では、 あくまで初心者向けの神経解剖学の解説としながらも、 こうした脳の栄養・代謝について取り上げた。 (が、いささかマニアックすぎたかもしれない。) 似たような用語が多数登場したため、 これを一気に覚えるのはたいへんかもしれない。 ぜひ何度か読みなおし、 それぞれの血管の名前と場所とあわせてイメージできるよう慣れてほしい。
コラム3: 血液脳関門
体内を循環する血液中には、 ニューロンの機能を阻害するようなホルモンや、 種々の化学物質がとけ込んでいる。 そのため神経系の組織は、 他の器官系と違い、 直接血液に暴露して栄養するわけにはいかない。 しかし一方で、 生きている以上、 神経系にも血液から栄養分や酸素を供給しなければならない。
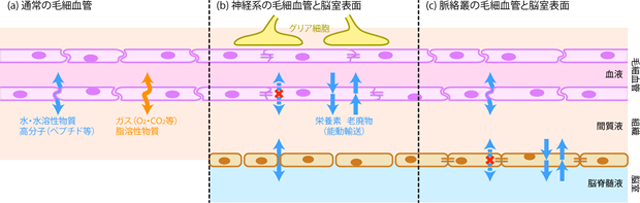
そこで神経系には、 神経組織に影響を与えるような化学物質は遮断しつつ、 必要な酸素・栄養素や、 いらなくなった老廃物だけを選択的に血液とやりとりできる仕組みが存在する。 これを血液脳関門 blood-brain barrierという。 血液脳関門の実体は、 毛細血管壁を構成する細胞がつくる強固な細胞間接着と、 細胞膜を隔てた選択的輸送である(Figure 25)。 「関門」というけったいな名前が付いているが、 なにか門扉となるような解剖構造が存在するわけではなく、 神経系を栄養する毛細血管全域が有する分子細胞学的機構の呼称である。
通常の毛細血管では、 血管壁は内皮細胞と呼ばれる細胞1層だけと非常に薄くなり、 その細胞のすきまから血漿が染み出して周囲の組織を栄養している(Figure 25a)。 しかし神経系では、 毛細血管の周囲にいるグリア細胞 glial cell が内皮細胞に向けて細胞間シグナル物質を分泌する。 このシグナルを受け取った内皮細胞は、 タイト結合と呼ばれる分子機構によって細胞膜を強く接着し合い、 血漿の漏れ出る隙をなくしてしまう。 こうして神経組織が血中成分に暴露されることを防いだうえで、 必要な物質のやりとりは、 内皮細胞の膜表面に存在するタンパクによる膜輸送によって行なうのだ(Figure 25b)。
このような仕組みにより、 代謝・循環における脳への物質の移行は厳しく制限されている。 そもそも脳が主たるエネルギーとしてグルコースしか使えないのも、 血液脳関門によって血中分子の直接的な移動ができないためである。 脂肪酸を筆頭とした他の栄養素は細胞膜を透過することができないので、 血液脳関門の存在下でも膜タンパクによって能動的に輸送されるグルコースに頼らざるを得ないのだ。 しかしそれと引き換えに、 神経系は血中の有害物質や感染源から物理的に守られる。 もし脳へと作用する薬物を開発しようと思ったら、 膜タンパク輸送の隙を突くなど、 なんらかの方法でこの障壁をすり抜けられる薬理物質をデザインする必要がある。 いくらGABA入りのチョコレートで神経伝達物質を経口摂取したところで、 それが血液脳関門を通過できなければ何の意味もない。
ちなみにちょっと前述したが、 脳脊髄液の産生に携わる脈絡叢においては、 脳室壁を構成する細胞が物質の行き来を制限している。 すなわち脈絡叢においては血液脳関門が存在せず、 毛細血管からは血漿成分が自由に浸み出せる。 (実際のところ、 脈絡叢の毛細血管は通常の毛細血管よりも物質透過性が高い。) そのかわり、 脈絡叢では脳室壁をつくる細胞同士がタイト結合によって強固に連結し、 血漿の拡散を防ぐとともに、 膜タンパクによる能動輸送などを行なう。 そのため結局のところ、 血液脳関門のない脈絡叢においても、 血中の化学物質が脳室内の脳脊髄液へと自由に拡散することはなく、 厳密な物質輸送の管理がなされている(Figure 25c)。
3.6 大脳(外観)
ここまで、脳を保護する骨や髄膜、 あるいは脳の栄養に関わる脳脊髄液や血液の循環について説明した。 以降では(やっとのことで)脳自体の解剖構造を解説していく。 本項ではまず、 認知神経科学においてもっとも重要な脳部位のひとつとなる大脳について、 その外観をみていこう。
大脳はヒトの脳のなかでもっとも大きい容積を占める部分である。 大脳は右脳と左脳という2つの 大脳半球 cerebral hemisphereからなる。 この半球を分けている深いミゾを 大脳縦裂 longitudinal fissureという。 左脳と右脳は機能的に違いがあるが、 解剖学的構造はほとんど変わらない。
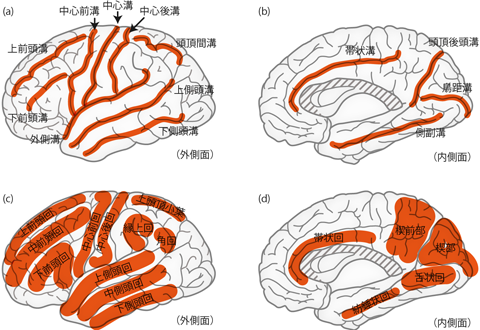
大脳の外観を眺めてみると、 ご存知のとおり、 大小たくさんのミゾが確認できる。 このような大脳の表面に存在するミゾを 溝 sulcus(こう)という。 また溝の中でも特に深いものは 裂 fissureと呼ばれることもある。 大脳を側面からみると、 とくに大きな2つの溝が確認できる(Figure 26)。 前後のちょうど真ん中あたりを頭頂部から腹側へ伸びているのが 中心溝 central sulcus (ローランド溝 fissure of Rolando)、 前方やや腹側から後方へ伸びて大脳を上下に分断しているのが 外側溝 lateral sulcus (シルビウス溝、 シルビウス裂 fissure of Sylvius)である。 外側溝はとくに深いので、 裂の名で呼ぶことも多い。 溝と裂には必ずしも厳密な呼び分けがあるわけではないが、 大脳の場合、 基本的には硬膜・クモ膜が入り込むような深いミゾを裂と呼ぶ。 例えば前述の大脳縦裂(左右大脳半球間のすきま)には硬膜・クモ膜が鎌状に入り込み、 これを大脳鎌と呼ぶのだった。
中心溝と外側溝により、 大脳半球は4つの部分に分けられる。中心溝の前方の部分を前頭葉 frontal lobe、 後方の部分を頭頂葉 parietal lobeという。 また外側溝の下の部分を側頭葉 temporal lobeと呼ぶ。 後頭葉 occipital lobe は大脳半球の後部を占める領域であるが、 後頭葉と頭頂葉・側頭葉とのあいだには、 外面観において明瞭な区切りとなる溝などはない。
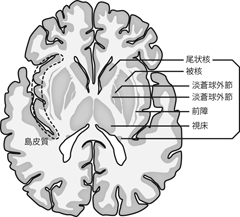
大脳の表面は外にみえている部分だけでなく、 大脳縦裂に埋もれた正中面の内側部にも広がっている(Figure 26d)。 また外側溝の内部にも、 外からはみえない埋もれた領域が存在する。 この領域は、脳標本において外側溝を上下に押し広げるように開くと観察できる。 あるいは次項で扱うように、 脳をスライスして前額断や水平断で確認してもわかりやすい(Figure 28)。 この外側溝の底に埋もれた領域を島 insula(とう) または島皮質 insular cortexという。 「島」はこの外側溝内部の領域の固有名詞であり、 溝のなかに隠された脳領域一般を指す名称などではない。
大脳には中心溝・外側溝の他にも、 大小たくさんの溝が存在する。 そして溝と溝のあいだは、 畑の畝のように盛りあがった凸形状をしている。 このような溝と溝で区切られた盛りあがりの部分を 回 gyrus(かい)という。 たとえば中心溝の前方の畝は中心前回と呼ばれる。 このように、 脳表面に走る溝とそれによって区切られた回には、 それぞれ名前が付けられている。 しかし溝と回の入り方の細部は個人差が大きいので、 解剖図説において、 細かな溝の分岐の仕方などまで闇雲に暗記することには意味がない。 Figure 26にヒトの大脳に一般的に観察される主要な回と溝をまとめた。 これらについては名前を聞いてすぐ場所をイメージできるよう、 また脳標本やMR画像をみてすぐに名前が挙げられるよう、 その位置関係をよく把握しておいてほしい。
3.7 大脳(断面と内部構造)
前節では大脳の外観について説明した。 本節では、 大脳の断面でみえる構造と、 内部に埋もれた神経組織について説明していこう。
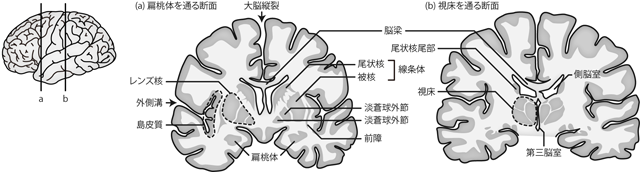
Figure 27は大脳の前額断(顔面と平行な断面で切った断面)である。 図から明らかなように、 大脳にも脊髄と同様、灰白質と白質がある。 灰白質はニューロンの細胞体がいる場所であり、 白質は軸索が通っている部分だ。 これは脳でも脊髄でもおなじである。 しかし思い出してみると、 脊髄の横断面では内部に蝶形の灰白質があり、 そのまわりに白質が存在した。 大脳ではその関係が逆転しており、 表面に灰白質が、 内部に白質がある。 脳において表面に露出したこのような灰白質を一般に皮質 cortexといい、 大脳の場合には大脳皮質 cerebral cortexという。
このように大脳においては、 表面の灰白質にニューロンが集まっており、 それらが連絡しあう軸索が白質として脳の内部に詰まっている。 ただし大脳に存在する灰白質領域は、 表面の皮質のみではない。 Figure 27やFigure 28にみられるとおり、 大脳には大脳皮質のほかに、 内部の白質に埋もれた灰白質のかたまりが存在する。 このように脳において、 白質のなかに埋もれて存在する灰白質を 核 nucleus(神経核、 皮質下核 subcortical nucleus)と呼ぶ。 灰白質はニューロンの細胞体のいる部分であるから、 核とはすなわち、 見渡す限り軸索の砂漠のなかに、 突如としてニューロンが集落をつくっているオアシス都市のような存在である。
脳内にはこのような皮質下の神経核が複数存在する。 たとえば側頭葉前端の内部には、 扁桃体 amygdala(amygdaloid body) と呼ばれる神経核がある。 扁桃とはアーモンドの実のことで、 扁桃体はその名のとおり、 アーモンドの実に似た歪んだ球形の神経核である。 扁桃体は情動機能への関与が知られる重要な神経核で、 動物における恐怖条件づけ学習などに深く関わるニューロンがみられたり、 ヒトの機能的脳イメージングでも感情に関連した脳活動が観察される。
扁桃体とおなじ前後軸のレベルの前額断でみると、 島皮質の直下にあたる白質内に、 厚みのある3重の層になった神経核が観察できる。 内側の2層を 淡蒼球 globus pallidusといい、 そのうち内側のほうを淡蒼球の内節 internal segment、 外側を外節 external segmentという。 淡蒼球に覆いかぶさるように最外層に存在するのが 線条体 striatumである。 線条体は勾玉をつぶしたような独特の形状をもち、 ひろい玉の部分を被殻 putamen、 被殻の背内側に沿うようにして存在する尾のような部分を 尾状核 caudate nucleusという。 尾状核は吻側では被殻と合流しているが、 尾側においては被殻から分離し、 側脳室に沿うような位置に細長く孤立している。 ただし発生学的にも機能的にも被殻と尾状核は共通性が高く、 前額断でみるとかなり離れているようにみえるものの、 両者は合わせて線条体というひとつの核と考えるべきである。 一方で尾状核をのぞいた被殻・淡蒼球外節・淡蒼球内節は、 ちょうど携帯端末の無線電波強度のマークのようなきれいな3層にみえるため、 あわせてレンズ核 lentiform nucleus (lenticular nucleus)と呼ばれることもある。 これはあくまで肉眼解剖において集合してみえる核のまとまりを命名したもので、 わざわざ被殻を尾状核から切り離したうえで、 淡蒼球とまとめて単一の核とみなす機能的な根拠はない。
線条体はこれらの皮質下核のいちばん外側に存在するが、 線条体と島皮質のあいだにはもうひとつ 前障 claustrum という核が存在する。 前障は非常に薄いものの、 大脳皮質の灰白質とは異なる神経発生によってつくられる皮質下の神経核である。
3.8 間脳
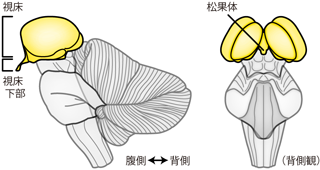
間脳はちょうど大脳に埋もれる位置に存在する脳領域で、 大脳皮質側からも脳幹側からも観察しづらい。 脳全体から大脳を取り除いたとすれば、 脳幹の突端に残ることになる2つのソラマメ型の構造が間脳である(Figure 29)。 ただし、 こうした仮想的なイラストはよく教科書等で見受けられるものの、 実際の間脳の表面には、 大脳との区切りになるような膜などは存在しない。 よって実物の脳標本において、 大脳だけをくるりと剥いてFigure 29のように間脳のみを剖出することはまず不可能である。 断面図でみた場合、 間脳は前述の皮質下核群と似て、 大脳の白質内に埋もれた灰白質のかたまりとして観察される。 しかしこれは脊髄・脳幹の灰白質の延長が大脳白質の基部に埋まっているのであり、 皮質下核のように全周囲を白質に取り囲まれているわけではない。 間脳は楕円球状の 視床 thalamusを主体とし、 その下方の視床下部 hypothalamus と上方の視床上部 epithalamus とに分類される。
視床にはさまざまな機能があるが、 その主たる役割は、 脊髄や脳幹から上行してきた感覚性線維の軸索を一時停車させ、 シナプスによって次のニューロンへとバトンを渡す中継地点となることである (視床中継核 thalamic relay nucleus)。 視床においてバトンを受け取った次の感覚性ニューロンは、 視覚なら後頭葉、 聴覚なら側頭葉上面など、 感覚種ごとに対応した大脳皮質領域へとさらに上行する軸索を伸ばす。 このとき異なる感覚種の中継を担うニューロン群は、 視床内のそれぞれ異なる決まった位置に集まっている。 このようなかたまりを別個の小さな核とみなすと、 視床は10以上の異なるニューロン群がつくる 視床亜核 thalamic subnucleusにより構成されていることになる (Figure 30)。 ただしこれらの亜核群は、 大脳の皮質下核のように明確な白質線維で分断されているわけではない。 組織学的にみると、 視床は全体としてやはり細胞体密度の高い灰白質であり、 そのなかで異なる機能をもったニューロンが棲みわけしているのを、 便宜上、核と呼んでいる。
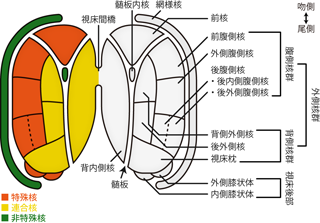
| 位置 | 名称 | 略称 | 種類 | 入力 | 出力 |
| 髄板の前方 | 前核 | 特殊 | 乳頭体・海馬・帯状回 | 大脳辺縁系 | |
| 髄板の外側 | |||||
| – 腹側核群 | 前腹側核 | VA | 特殊 | 黒質網様部・淡蒼球内節 | 運動野 |
| 外側腹側核 | VL | 特殊 | 小脳核・黒質網様部・淡蒼球内節 | 運動野 | |
| 後腹側核 | VP | ||||
| – 後外側腹側核 | VPL | 特殊 | 後索-内側毛帯系 | 体性感覚野 | |
| – 後内側腹側核 | VPM | 特殊 | 三叉神経脊髄路 | 体性感覚野 | |
| – 背側核群 | 背側外側核 | LD | 連合 | 海馬・帯状回との相互投射 | |
| 後外側核 | LP | 連合 | 視覚野・体性感覚野・頭頂連合野との相互投射 | ||
| 視床枕 | Pul | 連合 | 視覚野・体性感覚野・頭頂連合野との相互投射 | ||
| 髄板の内側 | 背内側核 | MD (DM) | 連合 | 前頭連合野との相互投射 | |
| 視床後部 | 外側膝状体 | LGN | 特殊 | 視索 | 一次視覚野 |
| 内側膝状体 | MGN | 特殊 | 下丘腕 | 一次聴覚野 | |
| 髄板内 | 髄板内核 | IL | 非特殊 | 痛覚伝導路・大脳辺縁系 | 皮質広範 |
| 視床外側表面 | 網様核 | TRN | 非特殊 | 他の視床核・大脳皮質 | 皮質広範 |
Table 3に主要な視床亜核とその機能をまとめた。 ただし現時点でこれをまる覚えしようとするのは、 単にたいへんというだけでなく、 本質的な理解をともなわないという点でも無駄が多い。 というのも感覚種ごとの機能論において、 各種感覚情報が受容器から大脳にいたるまでの経路を学ぶことになるが、 その際、 視床のそれぞれ異なる核が中継地点として登場する。 まずそれらの各論を学習すると、 機能的な視点での理解の縦糸を通すことができる。 そのうえで、 「それでは各感覚種で出てきた視床の核は、 視床のなかでどのように並んでいるのかな」 とTable 3に立ち戻ってみなおすことで、 視床という解剖構造を単位とした横糸の理解によって学習内容を整理することができる。 視床亜核群のはたらきは、 最初から一気に覚えようとするより、 このように順を追って勉強したほうが効率がよい。
さて、視床とともに間脳を構成する視床下部と視床上部についても簡単に触れておこう。 視床下部には、 情動や内臓機能に関連するさまざまなニューロンが存在し、 その機能に基づいて、 視床とおなじく多数の小核にわけられている。視床下部のおもな機能は、 情動行動の誘発や自律神経機能の調節、 体温調節、 概日リズムの調節などである。 また視床下部は血中にホルモンを分泌する内分泌器官である 下垂体 pituitary gland(脳下垂体) とつながっている。 このため視床下部は、 他の脳部位とニューロンによって神経連絡するだけでなく、 脳下垂体からのホルモン分泌を調節することで、 血液循環を介して全身の器官に影響を与えることができる。
視床上部には松果体 pineal gland(しょうかたい) という構造がある。 松果体には神経細胞も存在するが、 基本的に松果体は神経系の器官ではなく、 ホルモンを分泌する内分泌器官である。 視床上部の構造は松果体のみなので、 わざわざ視床上部といった分類をせずに 「間脳=視床+視床下部+松果体」と表現することもできる。 内分泌器官である松果体を脳に含めなければ、 もっと乱暴に「間脳=視床+視床下部」といってしまうことも可能である。 (実際、学会発表などでもよくみかける。) 松果体の細胞は、 概日リズム調節に関わるメラトニンというホルモンを合成・分泌する。 発生学的にみると、 松果体は脊椎動物の進化の過程で失われた頭頂眼という構造と同一の起源をもつ。 頭頂眼は文字通り「頭頂部の目」であり、 一部の爬虫類やヤツメウナギなどでは、 光を感知する第三の目として機能する。 体内の概日リズムをつかさどる内分泌腺が、 外界の昼夜を感知できる頭頂部の光受容器官を進化的起源とすることは興味深いが、 その因果関係については現在でもわかっていない。
先に名前は登場していたが、 この間脳のレベルに存在する脳室が第三脳室である。 第三脳室は左右の視床と視床下部にはさまれるかたちで、 わずかな薄い空洞を形成している。 いいかえれば、 左右の視床は第三脳室によって分断されている。 ただし第三脳室のなかほどには 視床間橋 interthalamic adhesion と呼ばれる点があり、 これがまるでピンポイントを接着剤でとめたように、 左右の視床を部分的につないでいる。 (個人差により視床間橋が存在しない場合もある。)
3.9 中脳
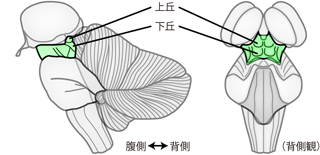
中脳は間脳と橋のあいだのごく小さな領域である。 外観では背側にある1対ずつの 上丘 superior colliculusと 下丘 inferior colliculus (あわせて四丘体 corpora quadrigeminaと呼ぶ) が特徴的である(Figure 31)。 小脳を下方に押し下げるようにして、 大脳と小脳のあいだから脳幹の背面をのぞきこむと、 ちょうど上丘と下丘が蚊に刺された跡のような4つのふくらみとして観察できる。 ヒトの場合、上丘は眼球運動や視覚情報の、下丘は聴覚情報の処理に関わっている。
中脳の内部には、 赤核 red nucleus・ 黒質 substantia nigraをはじめとして、 運動制御に関わる種々の核群が存在する。 また、中脳から延髄までの範囲にわたって、 脳幹の腹側には 脳幹網様体 reticular formation と呼ばれる構造が長く続いている。 網様体とはニューロンの軸索と細胞体が混ざり合った構造である。 脳幹網様体の内部には様々なニューロンが混在しているため、 その機能は一概にはまとめられないが、 中脳のレベルの網様体を刺激すると多様な不随意運動が生じることから、 とくに運動制御に関連したニューロン群が密集していることがわかる。
中脳のレベルでは脳室は非常に細くなっており、 第三脳室と第四脳室をつなぐ細い水道管の役割を果たしている。 そのためこの範囲の脳室を中脳水道と呼ぶ。
3.10 橋
橋は中脳と延髄のあいだにあり、 脳幹のなかでも明確に太く膨らんだ部分である(Figure 32)。 橋は両外側から出る線維によって小脳とつながっている。 この橋と小脳をつなぐ線維を 小脳脚 cerebellar peduncleと呼ぶ。 橋やその腹側の橋網様体には、 脳神経が担う感覚・運動双方の線維連絡の中継核が多数存在する。 また背後に控える小脳が大脳皮質や他の脳領域と連絡する際、 その改札口となる中継核も橋に存在している。
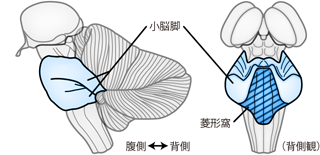
中脳のレベルでは細くせばまっていた脳室は、 橋に達すると左右にひろがって大きさを取り戻す。 これが第四脳室である(Figure 18)。 橋の背側には小脳が存在し、 小脳が小脳脚によって橋に抱きつくようなかたちとなっているため、 第四脳室はこのリア充カップルの背中(橋背面) とお腹(小脳腹側面)のあいだのすきまに相当する。 小脳脚を切り離し、 小脳を取り除いて脳幹を背側から観察すると、 第四脳室のスペースが脳幹背面の菱形のくぼみとして確認できる。 これを菱形窩 rhomboid fossaという。 ようするに第四脳室から小脳という「ふた」を外したとき、 脳幹側に残るくぼみが菱形窩である。
3.11 延髄
延髄は脳のうちもっとも尾方の部分で、 橋と脊髄のあいだに存在する(Figure 33)。 必然的に、 延髄は脊髄から脳へと上行するすべての線維と、 脳から脊髄へと下行するすべての線維の通り道になっている。 意識して考えないと、 脳の下端である延髄はなんとなく「首」というイメージを抱きがちだが、 実際の延髄は首というより頭のつけ根に位置する。 自分の後頭部を触りながら再確認してほしい。 脳は大脳から延髄まで含めて、 すっぽりと頭蓋腔内におさめられている。 頭部の背面をまわってちょうど両耳を結んだあたり、 後頭骨の隆起から頚部に向かってくぼみ落ちた部分を触って、 頭蓋骨の下縁を確かめよう。 その内側に小脳がおさめられており、 小脳の腹側に橋・延髄がある。 よって延髄は、 後頭部の隆起の下端、 頭から首へと移行するあたりに位置している。 ここより下では脊椎が中枢神経系を守る役割を担い、 その内部にあるのは頚髄よりはじまる脊髄である。
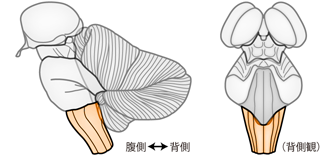
延髄には感覚性・運動性のおおくの神経核が存在する。 たとえば延髄の腹側には 下オリーブ核 inferior olivary nucleus という巨大な運動核がある。 オリーブ核はヒトにおいてよく発達しており、 外観からでも延髄腹側の特徴的な隆起として観察できる。また延髄背側には 薄束核 nucleus gracilis・ 楔状束核 nucleus cuneatus という体性感覚の中継核がある。 これらの運動・感覚核以外にも、 延髄には呼吸や血液循環など、 生命維持に不可欠な調節機能を担う核が集まっている。 そのため事故や疾病による延髄の損傷は、 致命的である場合が多い。
脳室についてみてみると、 橋と小脳にはさまれてできたすきまである第四脳室は、 だんだんと細くなりながら延髄のレベルまで続いている。 これは菱形窩におけるダイヤモンド型の下側の漏斗状の部分にあたる。 延髄は尾側で脊髄へとつながり、 それにともなって第四脳室も細い中心管へ移行する。
3.12 小脳
小脳は左右の小脳半球 cerebellar hemisphereと、 半球にはさまれた正中部の 虫部 vermisと呼ばれる隆起からなる(Figure 34)。 小脳の表面には非常に細い多数の小脳回 cerebellar foliaが存在する。 小脳を矢状断でみると、 樹枝状に広がる白質を灰白質が包んでおり、 まるで樹木のようにみえる。 これを小脳活樹 arbor vitaeと呼んでいる。
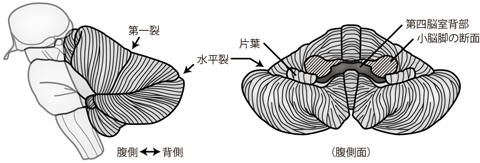
表面の小脳皮質には プルキンエ細胞 Purkinje cell という大きく特徴的なニューロンが存在する。 プルキンエ細胞は非常に多くの樹状突起を小脳皮質表面に向かって伸ばしている。 この樹状突起には、 感覚情報や運動情報を伝えるニューロンの軸索がシナプス結合する。 一方プルキンエ細胞の軸索は白質へと伸び、 小脳内部に存在する 小脳核 cerebellar nucleusへといたる。 ちなみにプルキンエ細胞の名前は発見者に由来したものだが、 「プルキンエ」という読みはチェコ語での発音を重んじた表記であり、 英語においては「ぷるきんじー」のように発音される。 よって外国人研究者との会話で「プルキンエセル」と言っても、 わかってもらえないことが多い。
小脳は感覚情報にもとづいて現在進行中の運動をモニタリングし、 自動的に運動を調節する機能をもっている。 小脳は、 大脳や脳幹によって企図された運動(理想)と、 その運動指令によって実際に生じた体の動き(現実)の両方の情報を受け取る。 そして両者の差を計算し、 その差を打ち消すような調節を次の運動指令に加える。 これにより実行される運動が、 もともと企図したものへと逐次是正されていく。 スポーツに熟達するにつれ、 はじめは意識的に苦労して行なっていた動作が次第に無意識にできるようになるのは、 小脳のはたらきのおかげである。 小脳が受け取る感覚情報の種類は、 固有感覚(筋・関節の位置感覚)・視覚・触覚・平衡核・聴覚など多岐に及んでいる。 運動を制御するうえで、 小脳はこれらの感覚情報を総合して、 身体のおかれている状況を判断するのである。
3.13 脳神経
末梢神経のうち、脳に出入りするものを脳神経といった。 いまさら勘違いするひとはさすがにいないだろうが、 「脳神経」と「脳の神経」はまったく別モノであるので改めて気をつけたい。 脳神経は、脳の腹側面から出入りする計12対の末梢神経線維である。 これらの線維には、 それぞれが関与する感覚・運動機能にもとづいた名称がつけられており、 同時に吻側から尾側の順に番号が振られている。 たとえば脳神経のうちもっとも吻側に位置する嗅神経は、 第I脳神経または脳神経Iと表わされる。 脳神経の番号にはローマ数字を用いるのが通例である。 各脳神経の主たる機能をTable 4にまとめた。
| 名称 | 種類 | 神経連絡 | 機能 | |
|---|---|---|---|---|
| I. | 嗅神経 | 特殊感覚 | 嗅上皮 | 嗅覚 |
| II. | 視神経 | 特殊感覚 | 網膜 | 視覚 |
| III. | 動眼神経 | 運動 | 外眼筋、内眼筋、上眼瞼挙筋 | 眼球運動 |
| IV. | 滑車神経 | 運動 | 外眼筋(上斜筋のみ) | 眼球運動 |
| V. | 三叉神経 | |||
| – 眼神経 | 感覚 | 眼窩、鼻腔、前頭部皮膚 | 体性感覚 | |
| – 上顎神経 | 感覚 | 上唇、歯肉、歯、頬 | 体性感覚 | |
| – 下顎神経 | 混合性 | 感覚枝: 歯肉、歯、口唇、舌 | 体性感覚 | |
| 運動枝: 咀嚼筋 | 咀嚼 | |||
| VI. | 外転神経 | 運動 | 外眼筋(外側直筋のみ) | 眼球運動 |
| VII. | 顔面神経 | 混合性 | 感覚枝: 味蕾 | 舌前2/3の味覚 |
| 運動枝: 表情筋、涙・顎下・舌下腺 | 顔面運動、唾液分泌 | |||
| VIII. | 内耳神経 | |||
| – 蝸牛神経 | 特殊感覚 | 蝸牛 | 聴覚 | |
| – 前庭神経 | 特殊感覚 | 前庭 | 平衡感覚 | |
| IX. | 舌咽神経 | 混合性 | 感覚枝: 味蕾、咽頭、頚動脈小体 | 舌後1/3の味覚、血流モニタ |
| 運動枝: 咽頭、耳下腺 | 嚥下、唾液分泌 | |||
| X. | 迷走神経 | 混合性 | 感覚枝: 咽頭、耳、鼓膜、臓器 | 咽頭の味覚、体性感覚、内臓覚 |
| 運動枝: 口蓋、咽頭、臓器 | 咽喉運動、副交感神経制御 | |||
| XI. | 副神経 | |||
| – 内枝 | 運動 | 口蓋、咽頭、喉頭 | 嚥下 | |
| – 外枝 | 運動 | 頚前外側の筋 | 頭部運動 | |
| XII. | 舌下神経 | 運動 | 舌 | 舌運動 |
脳神経のなかには、 単一の専門化した役割のみを担うものから、 さまざまな感覚・運動を手広く受けもつものまでさまざまがある。 そのすべてを一度に覚えるのは困難なので、 まずは各脳神経が求心性(感覚性)・遠心性(運動性)・ 混合性のいずれであるかを押さえておくのがよい。 というのも、 すべての髄節レベルにおいて必ず混合性神経だった脊髄神経と違い、 脳神経には混合性ではないものがあるからだ。 また一部の脊髄神経に内臓調節に関わる自律神経成分が含まれていたのとおなじく、 脳神経のなかにも自律神経性のものがある。 さらに感覚性の脳神経のなかでも、 視覚や聴覚など、 身体の一部のみに存在する特殊化した感覚器官で受容される感覚にたずさわるものは、 とくに特殊感覚性と呼ばれる。
ちなみに、脳神経の多様さは古くから医学部学生の悩みのタネであるらしい。 各脳神経の機能と線維走行の正確な把握は、 神経疾患などの臨床上、 重要な要暗記事項ばかりだからだ。 そこでせめて名前と番号くらいラクして覚えるために、 脳神経の覚えかたにはいろいろと語呂合わせが考案されているようだ。 もっとも有名なのは、 脳神経をIからXIIまで順に並べて文章にした、 「嗅いでみる、動く車の三つのそと、顔きく下に迷う副舌」 というものだろう。 (筆者も使っている。) まあ結局なんだか意味はわからないのだけど、 調子も軽快で口ずさみやすいので、 気にいったらこれで覚えてみるとよい。