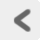@kanri_ninjin
@kanri_ninjin


 イクナイ! 581
イクナイ! 581







1 分類学の基礎
霊長類の分類の解説をはじめるに先立ち、 本節ではその基盤となる進化論や系統分類学の基礎知識を概説する。 多様なサルたちの生態についてはやく知りたい読者諸兄におかれては、 出ばなをくじく煩雑な用語説明など、うんざりだとおもう。 しかしこうした基礎知識は、 生物分類を学ぶうえでことあるごとに必要となる。 また多少とも進化論や分岐分類学について知っていると、 博物館や動物園にいったとき、 展示内容のみかたや理解の深さが変わってくるものだ。 本稿における専門用語や概念の導入は必要最低限のレベルにとどめるので、 一般教養をやしなうつもりで気楽に読んでいただけると幸いである。
1.1 リンネの階層分類
分類学 taxonomyとは、 世に数多存在する生物種たちを何らかの基準によって分類し、 人間にとって理解しやすいかたちに整理する学問である。 分類学を学問として確立したのは18世紀の植物学者 カール・リンネ Carl von Linnéとされている。

左: リンネの肖像画 右: 二名法による種の命名を提唱した『Systema Naturae(自然の体系)』(1735) [ウィキメディア・プロジェクトより転載(ともにPD)]
リンネは長く煩雑だった当時の生物種命名法に代わり二名法 binomial nomenclatureを導入した。 二名法において、種の学名は2語のラテン語で表わされる。 たとえばヒトは学名でホモ・サピエンス Homo sapiensだが、 このような2語からなる簡便な命名規則の発明はリンネの功績である。 彼はさらに、こうして命名した個々の種について、 似ているものをより大きな分類で順に包含していくことで、 世界に存在するすべての種を階層的に分類しようと試みた。 現在でも分類学では、 大きいほうから順に界–門–綱–目–科–属–種 (kingdom–phylum–class–order–family–genus–species) という階層的な分類を用いるが、 これはリンネとその後の分類学者たちによって考案されたものであり、 リンネ式階層分類 Linnaean hierarchyと呼ばれる。

筆者は高等学校の生物で、 大きいほうから「買い物項目家族で朱書き」と覚えた記憶がある。
これを用いると、たとえばニホンザルは「動物界–脊椎動物門33 正確には門のレベルでは脊索̇動物門とされ、 その下に脊椎̇動物亜門がおかれる。 ただ、脊索動物という単語はあまり一般的ではないので、 ここでは簡単のためこのような表現をとった。 –哺乳綱–霊長目–オナガザル科–マカク属–ニホンザル」と表わされる。 こうして「動物であり、さらに脊椎動物であり、さらに哺乳類であり…」というように、 大きいくくりから細かな区分へと段階的に分類していくことで、 その種が生物全体のなかでどういった立場に位置しているのかがわかりやすくなる。 これが階層分類の利点である。
1.2 進化論と系統学
リンネによって開かれた分類学は、 当時の混乱していた生物分類を体系的に整理し、 自然界の多様性に対する人間の理解を深めるのに役立った。 しかし一方で当時の分類学には、 分類の基準が研究者まかせであいまいであるという問題もあった。 たとえばリンネ自身は、 花の雄しべの数に基づいて植物を分類する方法を提唱した。 だが「なぜ雄しべの数を基準にすれば、 無数に存在する植物種を意味のあるグループにわけられる(はずな)のか」という妥当性に関しては、 必ずしも明白ではなかった。 実際、現代の分類学で遺伝的に近縁とわかっている種どうしでも、 雄しべの数が違っているものはたくさんある。 雄しべの数という客観的かつ定量的な特徴を分類の基準としたことは、 科学者たるリンネの姿勢として理にかなっているようにおもえる。 しかしそれが本当に生物の分類基準として適切なものであるかどうかは、別の問題なのだ。
このように、研究者ごとの分類基準と解釈如何によって結果ががらりと変わってしまうという問題に対し、 強固な「背骨」を通す役割を果たしたのが、 19世紀における進化論の台頭である。 進化論によれば、地球上のすべての種は、 変異と遺伝の繰り返しによって発生したはずである。 よって生物同士は、 「ある種から遺伝的変異によって別のこの種が生じた」という進化の道すじで結ばれる。 この道すじのことを、系統 phylogenyという。 系統学 phylogeneticsはそうした種間の系統関係を調べる学問であり、 結果として得られた系統関係を樹形図状に表わしたものを系統樹 phylogenetic treeという(Figure 3)。
一般に魚類と呼ばれている生物群は、 進化の過程で何回かにわけて独立に分岐したグループである。
系統樹は、原始的な祖先種から進化によって次々に種がわかれ、 最終的に個々の種にいたるまでの分岐の道のりを示す。 このため系統樹は分岐図 cladogramとも呼ばれる。
進化論に慣れ親しんだ現代のわれわれからすると、 このような系統関係による種の表現にどういった特別なメリットがあるのか、 なかなかわかりづらい。 しかし忘れてならないのは、 進化論と系統学が生まれる以前は、 世界中のすべての生物種は神が計画して創ったと考えるのが当然だったということだ。 科学者といえどもその例外ではない。 他ならぬリンネ自身、 分類学は、この世の生物群のありようから、 神による生命創造の計画を読み取る試みだと考えていた。 神によって創られたからこそ、 多種多様な生物のなかにも秩序だった分類が可能である、と。 しかし神の在不在は置いておくにしても、 科学者がそのような信念のもとに種を分類したとして、 それを神に「答え合わせ」してもらうすべがないことは明白である44 あたらしい分類をするたびにその答え合わせを神に尋ねていたら、 ロードス島の世界ではレベル10司祭の命がいくらあっても足りない。 。 よってそのような「神の意を読み取る」分類は、 結局のところ分類学者個人が「このようにわければ神の御心に沿う秩序だった分類になる」と (言い方は悪いが)勝̇手̇に̇感じた主観による分類になってしまう。
一方、進化論によれば、起源となった祖先種から、 進化によって現存の数々の種が生み出された進化の歴史(=系統)はたったひとつである。 系統学の利点は、 推定すべき「真の系統関係」が、 実際に過去の地球の歴史で起こった「唯一の史実」として規定されるところだ。 よって進化論を認めるならば、 分類学はそれぞれの学者のこころのなかに学者の数だけ存在する神の意図を追い求めるのではなく、 たったひとつ存在する真の系統関係を追究すればよい。 系統関係が明らかにできれば、 それはできあがった分岐図に含まれるすべての種を分類したのと同義である。 このように、進化の歴史である種間の系統関係を基準として生物種を分類する分類学を、 系統分類学 phylogenetic taxonomyという。
1.3 系統分類学の用語
系統分類学において種間の近縁関係の推定する方法を学ぶまえに、 用語の導入をしておこう。 Figure 4は、aからeの5種によって構成される生物群の系統樹を模式的に表わしたものである。

現生生物の系統樹では、葉は必ずおなじ高さにそろう。
系統樹は、もっとも原始的な単一の祖先種から、 問題としている個々の種までの進化の過程を樹形状に示したものだ。 このとき、木の「根元」の祖先種にあたる部分を根 root、 そこから分岐を繰り返してたどりついた木の「枝先」を葉 leafという。 根から葉にいたるまでには、 分岐点 nodeごとに枝 branchが2つに分岐していく。 最終的にたどりついた葉は、単一の種のこともあれば、 Figure 3のように生物群であることもある。
さて「分類」とは、 文字通り生物群をグループにわけることである。 これはFigure 4でいえば、 系統樹の葉である5つの種をどのようなグループにまとめるかという問題になる。 ここで系統分類では、 進化の歴史上の共通点をもつということを根拠に種のグループわけを行なう。 たとえばa種とb種は現在においては異なる種であるが、 進化の歴史を少し遡ると、 ひとつ前の分岐において合流している。 言いかたを変えれば、 a・b種はもともとは同一の種だったものが、 進化によって異なる種へと分岐したものと考えられる。 このように単一の祖先種から派生した子孫すべてを含む系統樹の部分集合を 単系統群 monophyletic group (分岐群 clade、クレード)という。 単系統群は系統分類学における種分類の基本単位である。 単系統群には、それに属する種数に制限はない。 よって前述のとおりa+bは単系統群だが、 たとえばa+b+cも、 a+b群とc種の分岐点以降のすべての種を含んでいるので同様に単系統群である。
一方、c+d+eというグループは単系統群ではない。 というのもこの場合、これらの種の共通祖先は、 もっとも古い分岐点である根そのものになる。 しかしc+d+eというグループは、 おなじく根の子孫であるa種とb種を含んでいないからだ。 このように、ある祖先から派生した種のうち一部を含まないため単系統群と呼べない部分集合を、 偽系統群 paraphyletic group (側系統群)という。 系統分類学において、偽系統群は意味のあるグループとはみとめられない。 というのも偽系統群は、単系統群から一部の種を恣意的に取り除いたものだからだ。 先の例を振り返ると、c・d・e種には 「根という共通の祖先を有している」ということ以外に系統樹から読み取れる類似性はない。 しかし「根から派生した」という性質は、a・b種も同様に有している。 ある生物群をグループとみなす根拠となった性質について、 同じくその性質を有する生物種が他にいるにも関わらずグループに含めないのは、 恣意的以外のなにものでもない。 よってこうした偽系統群は、 進化の歴史に基づいて生物種を分類するという場面においては、 根拠のない分類の仕方だといわざるをえない。 たとえばFigure 3をよくみると、 われわれが一般的に「魚類」と呼んでいる生物群は、 脊椎動物の進化の歴史のなかで独立に分岐したいくつかの枝の集合であることがわかる。 すなわち魚類は偽系統である。 もちろん「魚類」というくくりは日常生活上は便利だが、 厳密な系統関係に基づく分類という意味では不正確だということだ。
ところで種の分類を考えるにあたり、ある生物群の特徴は、 他の種との比較においてのみ浮き彫りになるものである。 よって系統樹を描く場合、 たいていは議論の対象である生物群だけでなく、 それに近縁な他の種までを含んだやや広い範囲の系統関係を記すことが多い。 このとき、系統樹のなかで興味の対象としている群を内群 ingroup、 それ以外の群を外群 outgroupと呼ぶことがある。 またある単系統群に対して、 その外群のなかでもっとも近しい群を姉妹群 sister groupという。
系統樹からは、ふたつの種のあいだの進化的な近さも読み取ることができる。 系統樹において根から葉に至る経路は、進化年代レベルでの時間経過を表わしていた。 葉に近い位置での分岐ほど、年代的に現在に近い時点で起こった種分岐である。 よって任意の2種について、 片方の葉からもう片方の葉へとたどる経路を考えたとき、 より根に近い分岐まで戻らなければならないとすれば、 それはその2種の進化的分岐が古い時点で起こったことを示している。 逆に2つの葉が、枝先に近い分岐ですぐ引き返すだけで辿れるなら、 それは2種の分岐が新しい年代で起こったことを意味している。 つまりその2種は、より近縁ということだ。 一方、系統樹においては、 葉どうしの作図上の近さは進化的な近さとは一致しない。 たとえばFigure 4において、 葉の位置の直線距離という意味では、 c種・d種間のほうがc種・e種間よりも近い。 しかしこれは、単にd・e種の分岐のあと、 d種が上に、e種が下に作図されたことによるものだ。 系統分岐上はd・e種のどちらについても、 根まで遡らないとc種にたどりつけないという意味でc種との近縁性は変わらない。
1.4 種の近縁性
系統分類学は分類の基準を唯一無二の進化の歴史にもとめる。 これにより分類学は、人為的な分類基準に依存しない、 より客観的な分類体系を構築することができる。 しかしそのさらなる利点は、真の系統がひとつしかないということだけではなく、 その系統を推定する際、 種間の近縁性の判断を客観的・論理的な方法でつけられることである。
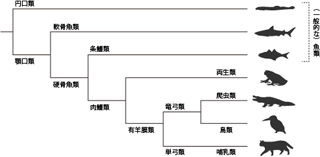
例として、見覚えのない架空の生物群における系統関係を考えてみよう(Figure 5)。
本図の作成にあたりインターネット検索をして、 はじめてはぐれメタルキングの存在を知った。 最近はこんなのまでいるんですね。
分類をするうえでは、 当然ながら対象の生物たちがもつさまざまな特徴を手がかりとする。 この特徴のことを、分類学では形質 traitと呼ぶ。 aからdの4種からなるこの生物群を観察してみると(Figure 5A)、 粘液質のからだという形質は全種に共通しているが、 そのほかにも一部の種のみに固有の形質もあるようだ。 たとえばb・c・d種はからだをメタル化させており、 しかもc・d種は涙型の形状を失って半液状化している。 そしてd種だけが、それらに加えて王冠をかぶるという特殊な形質を有している。
これらの形質を手がかりに、 種間の近縁関係を系統樹として表わすのが系統分類の目的となる。 そのうえでの前提は、 これらの形質が遺伝的変異によって生じたということだ。 つまりある形質は祖先種に偶然の変異として生じ、 それによって系統樹を「その形質をもつ種」と「もたない種」の2つの系統に分岐させる。 そのため系統樹においては、 各分岐点でわかれた枝の少なくとも一方に、 その分岐の根拠となった形質を記すことができる。 たとえばFigure 5Bは、 aからdの生物群について考えられる進化のシナリオのひとつを、 分岐の根拠となった形質とともに系統樹にしたものである。 もともと粘液質のからだだけをもっていた比較的単純な祖先種が存在し、 そこからまず、からだをメタル化させたものが別の種として分岐した。 さらにそのなかから半液状化したからだを獲得した種が分岐し、 その一部が冠をかぶってd種となった。 このような系統樹を想定した場合、 d種ともっとも近縁な種は、 最後の分岐においてd種とわかれたc種ということになる。
しかしこれら4種の生物群において考えることのできる進化のシナリオは、 これだけではない。 たとえばFigure 5Cは、別の進化の歴史の可能性を系統樹に表わしたものだ。 b種やc種が、祖先種からメタル化・半液状化を経て段階的に生じたと考える点は、 さきほどと変わらない。 しかしこの系統樹においては、 d種がb・c種とは独立にメタル化・半液状化・王冠という形質を獲得したと考えている点が、 大きく異なる。 この系統樹に基づけば、 d種ともっとも近縁なのはa種ということになる。
このように、ある一群の生物種からその系統関係を推測するにあたり、 考えることのできる分岐のシナリオは無数に存在する。 そこで拠り所となるのは、 新たな形質の獲得が偶然の変異によって生じたという進化論の前提だ。 生物のもつ形質はじつに多様だが、 それらは神の意志によって計画的に与えられたものではない。 遺伝子に生じた偶然の突然変異によってもたらされたものである。 よって、有用な形質が偶然生じる確率は、 非常に低いはずである。 ましてやそのような突然変異が、 別々の種で何度も独立に生じる可能性など著しく低い。 むしろ複数の種に共通している形質は、 それらの種が共有する祖先において突然変異によって一度だけ獲得され、 以降の子孫へは遺伝により伝えられていったと考えたほうがもっともらしい。 たとえばFigure 5Cの系統樹では、 d種の進化をb・c種とは別の枝と想定した結果、 系統樹全体でメタル化と半液状化の進化がそれぞれ独立に2回ずつ起こったことになっている。 またa種からd種が分岐するにあたっては、 3つもの形質が同時に獲得されなければならなかった。 このような「ぜいたく」な新形質の獲得は、 進化論においては考えにくい。 よって系統分類学では、 Figure 5Bの系統樹のように、 新しい形質が進化によって獲得された回数がより少なくてすむ説明を採用するのである。 これを最節約原理 principle of maximum parsimony (最節約法 maximum parsimony method)という。 最節約原理にのっとれば、 任意のデータについて考えられる無数の系統樹から、 客観的な基準でもっともらしいものを選択することができる。 この点において系統分類は、 それ以前の恣意的な分類学に勝っている。 これは進化論という理論的な基盤をもった系統分類学の利点といえる。
1.5 階層分類と系統分類
このように系統分類学は、 進化のメカニズムに照らし合わせた妥当性という観点により、 無数に考えられる可能性のなかからもっともらしい系統関係を推定する。 こうしてつくりだされた系統樹は、 分岐点ごとに形質に基づいて2つの単系統がわかれていく。 個々の種は、系統樹の根から葉にたどり着くまでの過程で、 どのような形質を根拠にどういった系統群からわけられてきたかによって特徴づけられる。 こうした分岐図においては、ある意味ですべての分岐が同様に重要である。
このような系統に基づく分類は、進化論がひろく認められた現在における分類学の基本となっている。 しかしその一方、生物分類においては、 古典的なリンネ式階層分類の表現も今なお並行して使われ続けている。 たとえば新しい動物について話題に出すときは、 それが「なに目・なに科・なに属」なのかを明示することがおおい。 というのも、界–門–綱–目–科–属–種という階層的な分類は、 分類全体のなかでの種の「位置」を指し示すには非常に便利なのだ。 「Saimiriの特徴」といわれてもその単語を知らなければどうしようもないが、 「Saimiri属の特徴」といわれれば、 少なくとも属のレベルでの比較的詳細な性質について議論していることはわかるだろう55 Saimiriはリスザル属のこと。 。 ちなみに、あまり生物分類に詳しくないひとは、 いわゆる「哺乳類」というのが綱のレベル (哺乳綱 Mammalia)の分類群であることを覚えておくと、 階層分類におけるだいたいの感覚をつかみやすい。 また本稿ではサルの仲間の分類を扱うわけだが、 いわゆる「霊長類」は階層分類における目のレベル (霊長目)に相当する。 よって次節以降では、 基本的に目以下の細かい階層についての分類をしていくことになる。
系統分類学において系統樹を描くにあたっても、 リンネ式階層分類の表現の併用は非常に一般的だ。
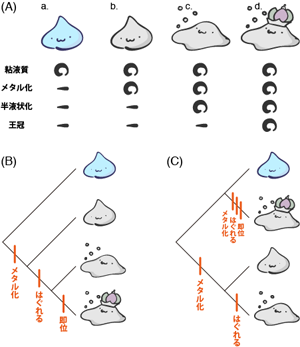
階層分類上おなじレベルの系統群が次々に分岐するとき(A)、 これらがおなじ分類段階であることを強調しようとすると、 系統樹は三つ叉以上に分岐する(B)。
たとえばFigure 6Aはオナガザルの仲間の系統関係を示している。 この系統樹では、葉は個々の種にいたるまで詳細には描かれておらず、 「なになに属」というレベルで終わっている。 本当の意味での系統分類には「どの時点での分岐が科のレベルで、 どこからが属」というような階層構造はないため、 階層分類の表現を系統樹のなかにもちこむ必然性はない。 実際、古典的階層分類におけるレベルについて言及したくない場合には、 「なになに類」や「なになにの仲間」という表現も頻繁に用いられる。 しかし系統樹においても、 「いわゆる階層分類的な区分でいくと属のレベルであるマカクの仲間」 という意味で「マカカ属」と表記しておけば、 それがどの程度の広さでの分類区分なのかがわかりやすい。
もちろん、系統分類と階層分類は本質的には両立しないものだから、 さまざまな部分で齟齬が生じる。 たとえば系統樹は各分岐点で枝が2つにわかれる二分木だが、 界–門–綱–目–科–属–種という階層は7段階しかない。 だから大きいほうから順に生物を2つずつにわけていったのでは、 最終的に葉にたどりつくまでに27 = 128種類にしか分岐できず、 全生物種を分類するには到底たりない。
こうした階層分類の併用による問題は、 さまざまなやりかたで柔軟に対処される。 そもそもリンネ式分類における階層は、基本となる7階層に加え、 各段階に上 super-・ 亜 sub-・下 infra-といった接頭辞をつけることで、 より細かな階層を派生させることができる。 たとえば門よりは小さなくくりだが、綱よりはひろい範囲の生物群を命名する場合、 亜門subphylumや上綱superclassといった階層をつかう(e.g., 脊椎動物亜門)。 また系統樹においては、 分岐ごとに階層分類的な段階が進むとは限らない。 たとえばFigure 6Aに示したとおり、 オナガザル亜科のなかでは、 マカカ属・マンガベイ属・ヒヒ属といった属のレベルの分類群が順々に分岐していく。 系統樹において、ある分岐で「属」を分岐させたから、 その次の分岐では1階層下の「種」を分岐させなければならないという決まりはないのだ。 異なる時点において分岐したグループたちが同じ階層レベルの分類群とみなされるのは、 一見奇妙かもしれない。 しかし、実際の進化は連続的な分岐により起きており、 階層分類はそれを恣意的にいくつかの段階にわけて分類しているだけだと理解していれば、 同一レベルの分類群が順々に分岐することにも疑問は生じないだろう。
これと関係するが、 一般に系統樹における分岐は、必ずしも二分岐のみとは限らない。 ひとつの分岐で3つ以上の枝がわかれる分岐図も、 現実には頻繁に目にする。 これにはふたつの理由が考えられる。 ひとつは先ほど説明したように、 階層分類においてはおなじレベルに区分される分類群が、 次々と分岐しているような場合である。 これらの分類群が、 系統樹のなかでどういう順に派生したのかにはとくに興味がなく、 むしろそれらが階層分類的におなじ段階として扱われることを強調したい場合、 Figure 6Bのように一括した多重分岐として描く場合がある。 しかし一方で、いくつかの分類群の分岐の順が本当にわからないこともある。 実際の進化は複数の系統が順々に分岐することによって起こったのかもしれないが、 現実に調べられる特徴においては、 すべての群がおなじ程度似ているという場合だ。 こういうときには、やむを得ないので、 3つ以上の分類群が一度に分岐する系統樹を描くことになる66 もちろん、進化の歴史のなかで、 本当に3つの生物群への分岐が同時に生じたという可能性もある。 たとえばあるとき起こった地殻変動により、ひとつの種が3つの地域に同時にわけ隔てられ、 それらが独自の進化を遂げた場合などだ。 このようなケースでは、真の系統樹の形が三分岐以上ということになろう。 しかし現実的には、 3つ以上の分類群が同時的に分岐したという積極的な証拠があることはあまりない。 系統樹において三分岐以上の分岐が表われたら、 たいていは本文中に記したような理由によるものだ。 。