@kanri_ninjin
@kanri_ninjin


 イクナイ! 581
イクナイ! 581







第36回 日本神経科学大会(2013)の記録
2013年の神経科学学会に参加した際の記録です。見聞きしたことを忘れないための、至極個人的なメモです。
理解不足や間違え等もあるかとおもいますので、悪しからず。
発表者が複数いる場合、ファーストとラスト、あるいはラストに準ずるひと2~3人だけ表記します。
(みやすさの優先のため。)
アスタリスクつきのポスターは、自分でちょろっと読んだだけのヤツ。
アスタリスクなしは、ちゃんと発表者に説明してもらったヤツです。
基本情報
今年は3年に一度の3学会合同大会。会場は京都国際会館。
なので通勤はフツーにバスで。
神経科学大会としての大会長はBSIの加藤忠史先生。
去年はプログラム冊子まで会場で渡されるケチ仕様でしたが。
今年は合同大会なためか、冊子は事前送付。
しかもネームカードも印字済みという太っ腹。
ぶっちゃけそんぐれーまいとsh(ry
あとプログラム厚い。
重い。
腰にくる。
(´д`)
それから、前年・前々年は会期が4日間で。
無駄にヒマな時間が多かったですが。
今回は合同大会にも関わらず、あくまで会期は3日間。
それゆえ、なかなか詰まった大会でした。
まあなんか、いちおう正式には会期は20日から23日までの4日間なんですが。
4日目の日曜日は、2会場しか使われてなくて。
なんか記念シンポと、脳科学教育コースとかいう謎な講義をやってるみたい。
なので大会としては、実質3日間です。
なんせアンケートにさえ、
「大会の長さはどれくらいがよいですか(本年は3日)」
って書いてあったからなwww学会側としても、4日目はものの数ではない認識なのだろう。
あと今年は、(少なくともあたしが参加した)JNSでは初の、午前・午後ポスター貼り替え制。
なので、チェックしたいポスターをちゃんと聞くのが、かなり大変でした。
まあ、そのおかげで会期3日におさまったんだろうけど。
これについての感想は、またページ最後で。
スケジュール
3日間の自分の行動。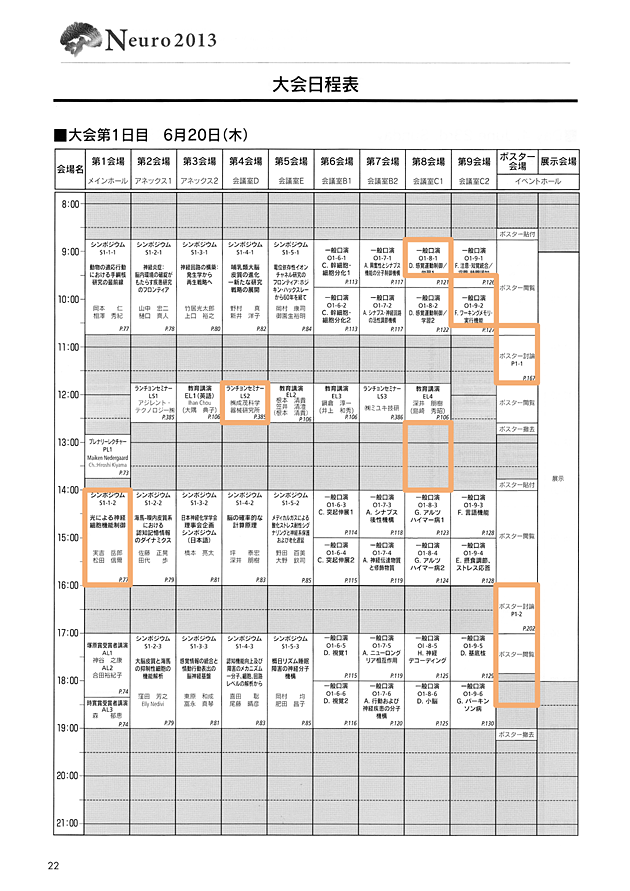


トーク
1日目(2013.06.20)
O1-8-1 感覚運動制御/学習1北澤 茂, 吉田 正俊
しょっぱなにしてまさかのオーラル。
いやなんか、初日午前はあまりひかれるシンポがなくてな。
1人目の喜多村さんは、ADHDにおけるサッケードRTのはなし。
gap effectを使うと、ADHD群のサッケード異常をよりうまく抽出できる。
ただし、そもそものRTの分散が大きかったりするようなので。
定量指標が、ADHD群におけるRTのどういう特性をとらえているのかは注意が必要っぽい。
2人目の小林先生は、マイクロサッケード。
アンチサッケード準備中のマイクロサッケード頻度から、その生物学的基盤を考える、と。
瞬目とかマイクロサッケードって、そもそもは被験者が「うっかりやってしまう」もので。
古典的な認知研究なんかにおいては、ある意味ノイズというか。
ジャマもの扱いな場合も多いが。
いい加減、眼球運動研究は、そのへんにいかないとネタ切れ気味ってことなのだろうか…。
3人目の大畑さんは、MEGをつかった運動デコード。
ただ、運動直前だったら、運動野からわりと正確なデコードできて当然なので。
Goシグナルが出るより前の脳活動からRTを予測してみようってトコが面白かった。
O1-9-1 注意・知覚統合/空間・時間認知いやなんか、初日午前はあまりひかれるシンポがなくてな。
1人目の喜多村さんは、ADHDにおけるサッケードRTのはなし。
gap effectを使うと、ADHD群のサッケード異常をよりうまく抽出できる。
ただし、そもそものRTの分散が大きかったりするようなので。
定量指標が、ADHD群におけるRTのどういう特性をとらえているのかは注意が必要っぽい。
2人目の小林先生は、マイクロサッケード。
アンチサッケード準備中のマイクロサッケード頻度から、その生物学的基盤を考える、と。
瞬目とかマイクロサッケードって、そもそもは被験者が「うっかりやってしまう」もので。
古典的な認知研究なんかにおいては、ある意味ノイズというか。
ジャマもの扱いな場合も多いが。
いい加減、眼球運動研究は、そのへんにいかないとネタ切れ気味ってことなのだろうか…。
3人目の大畑さんは、MEGをつかった運動デコード。
ただ、運動直前だったら、運動野からわりと正確なデコードできて当然なので。
Goシグナルが出るより前の脳活動からRTを予測してみようってトコが面白かった。
山口 陽子, 神作 憲司
前オーラルの4人目を聞かずに退出し。
となりの会場のオーラルに移動。
こちらの4人目のひとの発表を聞くことに。
…したはいいが、内容はサッパリでした。
まああたし自身、研究でやってる実験内容に興味があるというよりは。
解析手法のほうを知りたかっただけなので。
よしとする。
O1-9-2 ワーキングメモリ・実行機能となりの会場のオーラルに移動。
こちらの4人目のひとの発表を聞くことに。
…したはいいが、内容はサッパリでした。
まああたし自身、研究でやってる実験内容に興味があるというよりは。
解析手法のほうを知りたかっただけなので。
よしとする。
田中 啓治, 宮川 剛
引き続き同会場で、次のオーラルを聞く。
1人目は苧坂先生のところのtDCSの研究。
リーディングスパンで、ターゲット語が意味的に重要/非重要な2条件を使用し。
トップダウンの注意により、非ターゲット語からの干渉を抑える要求をつくる。
で、トップダウン注意に関わるといわれる頭頂皮質にtDCSを適用して効果をみた、と。
結果はなんとなく面白そうな印象だったんですが。
なんせ15分の時間制限に内容が詰め込まれているんで。
途中で電流刺激の効果のグラフを読み取りきれず、ロストしましたorz
てかtDCSって、アノーダルとカソーダルとシャムがでてきて、グラフ読むのに手間取ってしまう。
ワーキングメモリが足りないwww
2人目はJohn Duncanのところの楠先生。
じつのところ、こないだラボのJCでこきおろした(笑)、Neuronの論文の内容です。
そしてやっぱり、こういう解析をする理由がわからん。
百歩譲って、ユニット記録データを擬似的にポピュレーションにするのは認めても。
state spaceとユークリッド距離とMDSは、みてるもの同じなので。
ひとつの論文内で、本質的に同じ解析を3回も繰り返す意味がわからないし。
後半のデコーディング云々ってやつも、単純にデータを分けて相関とってるだけだから。
デコーディングじゃないよコレ。
ニューロン活動としては、ユニークなものが色々取れてる研究なはず。
なので、データにいちゃもんつけるつもりはないですが。
解析の仕方にかなり異議あり、というか。
たぶんこの解析に直接手をくだしたヤツ(楠先生ではない)は、
3人目の渡邉さんは、サルに二重課題をやらせて、課題干渉をニューロンレベルでみるという。
実験者およびサル双方の忍耐力を試す実験だよこれ。
課題負荷のスイッチにより、ニューロンの表現する情報も変わるのが綺麗。
認知容量の再配分が、課題関連ニューロンの活動として観察できるという。
こういうのは分散説明率より情報量でプロットするほうがよくみる気もするけど。
ちなみに4人目のかたは、ちょっとあたしの研究と関係なさそうだったので。
すたこら退出して、ポスター会場へ。
LS2 新皮質における回路とその機能1人目は苧坂先生のところのtDCSの研究。
リーディングスパンで、ターゲット語が意味的に重要/非重要な2条件を使用し。
トップダウンの注意により、非ターゲット語からの干渉を抑える要求をつくる。
で、トップダウン注意に関わるといわれる頭頂皮質にtDCSを適用して効果をみた、と。
結果はなんとなく面白そうな印象だったんですが。
なんせ15分の時間制限に内容が詰め込まれているんで。
途中で電流刺激の効果のグラフを読み取りきれず、ロストしましたorz
てかtDCSって、アノーダルとカソーダルとシャムがでてきて、グラフ読むのに手間取ってしまう。
ワーキングメモリが足りないwww
2人目はJohn Duncanのところの楠先生。
じつのところ、こないだラボのJCでこきおろした(笑)、Neuronの論文の内容です。
そしてやっぱり、こういう解析をする理由がわからん。
百歩譲って、ユニット記録データを擬似的にポピュレーションにするのは認めても。
state spaceとユークリッド距離とMDSは、みてるもの同じなので。
ひとつの論文内で、本質的に同じ解析を3回も繰り返す意味がわからないし。
後半のデコーディング云々ってやつも、単純にデータを分けて相関とってるだけだから。
デコーディングじゃないよコレ。
ニューロン活動としては、ユニークなものが色々取れてる研究なはず。
なので、データにいちゃもんつけるつもりはないですが。
解析の仕方にかなり異議あり、というか。
たぶんこの解析に直接手をくだしたヤツ(楠先生ではない)は、
「ウブな実験屋ごとき、この程度なインチキ手法でだまくらかせる」
と確信犯でやってるんでしょうね。3人目の渡邉さんは、サルに二重課題をやらせて、課題干渉をニューロンレベルでみるという。
実験者およびサル双方の忍耐力を試す実験だよこれ。
課題負荷のスイッチにより、ニューロンの表現する情報も変わるのが綺麗。
認知容量の再配分が、課題関連ニューロンの活動として観察できるという。
こういうのは分散説明率より情報量でプロットするほうがよくみる気もするけど。
ちなみに4人目のかたは、ちょっとあたしの研究と関係なさそうだったので。
すたこら退出して、ポスター会場へ。
村山 正宜
やはり初日は、いろいろ選択肢あっても、結局ナリシゲのランチョンになるよねっていう。
内容も豪華だし。
今回の演者は吉村先生と村山先生で。
吉村先生はいつもどおり、スライスで刺激マップやマルチパッチのかたーいデータ。
細胞接着因子を遺伝的に操作して、視覚皮質で細胞間結合確率もとめたり。
いっぽう村山先生は、運動皮質。
後肢を研究対象にするあたりが、ニッチだよね。
そしてさすが実績主義の理研BSIというか。
樹状突起イメージングから仮想オープンフィールドから。
とにかく手の広げかたがすごい。
vivoの全自動マルチパッチシステムとか、狂気の沙汰だとおもいました。
ナリシゲ脅威の技術力。
S1-1-2 光による神経細胞機能制御内容も豪華だし。
今回の演者は吉村先生と村山先生で。
吉村先生はいつもどおり、スライスで刺激マップやマルチパッチのかたーいデータ。
細胞接着因子を遺伝的に操作して、視覚皮質で細胞間結合確率もとめたり。
いっぽう村山先生は、運動皮質。
後肢を研究対象にするあたりが、ニッチだよね。
そしてさすが実績主義の理研BSIというか。
樹状突起イメージングから仮想オープンフィールドから。
とにかく手の広げかたがすごい。
vivoの全自動マルチパッチシステムとか、狂気の沙汰だとおもいました。
ナリシゲ脅威の技術力。
実吉 岳郎, 松田 信爾
今大会、メインホールの講演で唯一聞いたシンポ。
会場がでかすぎてびびる。
じつのところ、内容自体は自分にはあまり関係ないので。
込み入った考察をできる知識もないのですが。
スパインの機能形態解析がこんなにも「あたりまえ」なことに素直に驚きます。
このセッションでは、おもにCaMKIIがらみのアプローチが多かったですが。
LTP・LTDの生物学的基盤が、こんなミクロなレベルで理解できる時代というのは。
やはり二光子技術は偉大だということですよ。
会場がでかすぎてびびる。
じつのところ、内容自体は自分にはあまり関係ないので。
込み入った考察をできる知識もないのですが。
スパインの機能形態解析がこんなにも「あたりまえ」なことに素直に驚きます。
このセッションでは、おもにCaMKIIがらみのアプローチが多かったですが。
LTP・LTDの生物学的基盤が、こんなミクロなレベルで理解できる時代というのは。
やはり二光子技術は偉大だということですよ。
2日目(2013.06.21)
S2-2-1 大脳皮質回路の動作原理解明への最前線細谷 俊彦, 谷口 弘樹
2日目の午前は、皮質回路のシンポへ。
ぶっちゃけ5演題中3つは、サテライトで聞いたトークとほぼ同じ。
のこりふたつのうち1人目のYang Danは、大脳基底部を光刺激するハナシで。
コリン性ニューロンの刺激でV1ニューロンの活動や情報量が上がり。
動物の弁別成績もあがるという。
発想としては古典的で単純な脳幹網様体賦活系のイメージなんだけど。
それを光遺伝学などを駆使して、細胞の情報表現レベルで調べてるのがクール。
もうひとかたは、川口先生で。
皮質錐体細胞の分類のハナシだったんだけど。
中盤ぐらいでロストしました(笑)
細胞形態に加え、電気生理・結合・遺伝特性のデータが次々出てくるので。
どの細胞がどういうのだったか、途中からついていけなくなった。
論文のほうを参照して、じっくり復習します。
AL4 Dreaming of Mathematical Neuroscience for Half a Centuryぶっちゃけ5演題中3つは、サテライトで聞いたトークとほぼ同じ。
のこりふたつのうち1人目のYang Danは、大脳基底部を光刺激するハナシで。
コリン性ニューロンの刺激でV1ニューロンの活動や情報量が上がり。
動物の弁別成績もあがるという。
発想としては古典的で単純な脳幹網様体賦活系のイメージなんだけど。
それを光遺伝学などを駆使して、細胞の情報表現レベルで調べてるのがクール。
もうひとかたは、川口先生で。
皮質錐体細胞の分類のハナシだったんだけど。
中盤ぐらいでロストしました(笑)
細胞形態に加え、電気生理・結合・遺伝特性のデータが次々出てくるので。
どの細胞がどういうのだったか、途中からついていけなくなった。
論文のほうを参照して、じっくり復習します。
甘利 俊一
甘利先生が、昨年、文化功労者に顕彰されたということで。
その記念講演。
そりゃもう、ランチョン食いにいってる場合じゃねえっていう。
いやーやっぱ甘利先生のおはなしは面白いな。
発表中、何回"simple"って言ったか数えとけばよかった(笑)
あと、まさかのiPadプレゼンwwwww
でも、途中でスライドがスリープになっちゃって、
それはむしろすごいっていう。
その記念講演。
そりゃもう、ランチョン食いにいってる場合じゃねえっていう。
いやーやっぱ甘利先生のおはなしは面白いな。
発表中、何回"simple"って言ったか数えとけばよかった(笑)
「神が造ったなら、この世界はもっとunderstandableなハズ」
「でも実際は、世界はlaw of informationとlaw of mindで淘汰されてきた」
「それゆえ(私はsimpleなことが好きなのに)世の中complexなことばっかり」
「そのmessを理解するため、しょーがないからsimple mathematical modelを使う」
生粋の数学思考だ。「でも実際は、世界はlaw of informationとlaw of mindで淘汰されてきた」
「それゆえ(私はsimpleなことが好きなのに)世の中complexなことばっかり」
「そのmessを理解するため、しょーがないからsimple mathematical modelを使う」
あと、まさかのiPadプレゼンwwwww
でも、途中でスライドがスリープになっちゃって、
"This is the 2nd time I use this device."
"I don't know anything about computers."
っておっしゃってた。"I don't know anything about computers."
それはむしろすごいっていう。
ポスター
1日目(2013.06.20)
P1-1-82 ラットの吻側・尾側前肢運動野における運動発現と出力制御に関連したマルチニューロン活動齊木 愛希子, 木村 梨絵, 礒村 宜和
Go/No-goのデータどうなったかとおもって聞きにいったら、違う課題になってた…。
頭固定のマウスのレバー引き課題で、レバーの重さを2条件つくってRFA・CFAの活動とるっていう。
まあ結果としては、heavy選択性細胞もlight選択性細胞もいて。
heavyのほうが集団全体として活動があがる、みたいなことはいまのトコみられてない。
領域間差異もそれほどはない。
で、いいのかな…?
やっぱりGo/No-go課題のほうが気になるので、ついついそっちも質問してしまったけれども。
なかなかうまいことまとまらない、ってカンジなんでしょうか。
あ、ただ、なんかこのM2の定義が微妙らしく。
ここでのRFAは電気刺激で前肢運動がおこるエリアだけど。
川口先生とかは、むしろ運動誘発のおこらない領域をM2と呼んでたりして。
けっこう研究者によって違いがあるのだとか。
まーだとすれば、あたしだったらRFAにこだわらず内側から取るけどな。
P1-1-88 外発性・内発性運動に伴うラット一次・二次運動野の協調的神経活動*頭固定のマウスのレバー引き課題で、レバーの重さを2条件つくってRFA・CFAの活動とるっていう。
まあ結果としては、heavy選択性細胞もlight選択性細胞もいて。
heavyのほうが集団全体として活動があがる、みたいなことはいまのトコみられてない。
領域間差異もそれほどはない。
で、いいのかな…?
やっぱりGo/No-go課題のほうが気になるので、ついついそっちも質問してしまったけれども。
なかなかうまいことまとまらない、ってカンジなんでしょうか。
「齧歯類だとM2が意思決定情報もってる」
ってハナシだし、行動選択の余地がある課題のほうが、RFAならではの傾向でやすい気がするが。あ、ただ、なんかこのM2の定義が微妙らしく。
ここでのRFAは電気刺激で前肢運動がおこるエリアだけど。
川口先生とかは、むしろ運動誘発のおこらない領域をM2と呼んでたりして。
けっこう研究者によって違いがあるのだとか。
まーだとすれば、あたしだったらRFAにこだわらず内側から取るけどな。
木村 梨絵, 齊木 愛希子, 礒村 宜和
延々とひとだかりができてて、ちゃんと聞けなかった。
internalのハナシは自分の研究と関連してるので気になってるんですが…。
ただ、今回は課題関連活動そのものより、細胞間の関係に焦点をあててるっぽかったが。
遠巻きに読んだカンジでは、かなり解析苦しそうな印象が。
cross correlationからschematic diagram描いて結合かぞえるとか。
偶然とれた局所データにすごい依存しそうだけど、大丈夫なのか…?
P1-1-93 ヒトの注視行動に影響を与える要因:自然風景に物体画像を配置した複雑な刺激画像による解析internalのハナシは自分の研究と関連してるので気になってるんですが…。
ただ、今回は課題関連活動そのものより、細胞間の関係に焦点をあててるっぽかったが。
遠巻きに読んだカンジでは、かなり解析苦しそうな印象が。
cross correlationからschematic diagram描いて結合かぞえるとか。
偶然とれた局所データにすごい依存しそうだけど、大丈夫なのか…?
鈴木 実佳, 藤田 一郎, 田村 弘
いろんな背景画像のうえに、ターゲットとなるオブジェクトを表示し。
その対象の「注視されやすさ」を調べるという実験。
重なった領域のRGB値コントラストだけで予測できるのは、たしかに便利かも。
計算簡単だし。
はなしを聞いてたら、やはりサルのほうで背景+オブジェクトで実験してるので。
ヒトでも似たような課題構成つかって、行動でなんかできないか、という動機らしい。
…が、そのへんはあんま書けんわな。
P1-1-102 均一な方位選択性分布を示すニューロン集団の記録に適した多電極アレイのデザイン*その対象の「注視されやすさ」を調べるという実験。
「そんなもん、saliencyマップ計算すればよくね?」
と思わないでもないけど。重なった領域のRGB値コントラストだけで予測できるのは、たしかに便利かも。
計算簡単だし。
はなしを聞いてたら、やはりサルのほうで背景+オブジェクトで実験してるので。
ヒトでも似たような課題構成つかって、行動でなんかできないか、という動機らしい。
…が、そのへんはあんま書けんわな。
圓山 由子, 伊藤 浩之
2種の多点電極をネコ視覚野に刺して。
どちらのほうが、よりまんべんない方位選択性ニューロンをとれるかっていう。
予測部分の定式化とか、面白そうだったんだけど、ちゃんと聞く時間がなかった。
P1-2-75 ラットのリズムに基づく行動中または時間間隔に基づく行動中に見られる小脳活動*どちらのほうが、よりまんべんない方位選択性ニューロンをとれるかっていう。
予測部分の定式化とか、面白そうだったんだけど、ちゃんと聞く時間がなかった。
山口 健治, 高橋 晋, 櫻井 芳雄
fixed interのスケジュールをいじって、その間の小脳プルキンエ細胞活動をとったと。
聞きたかったが、時間が足りず。
たしか去年は
無事取れてるんだろうか。
P1-2-99 即時的な行動選択に関わる淡蒼球外節の神経活動聞きたかったが、時間が足りず。
たしか去年は
「プルキンエ細胞すごい取りにくい」
みたいなことおっしゃてたと思ったが。無事取れてるんだろうか。
吉田 篤司, 田中 真樹
pro-saccadeとanti-saccadeって、普通は注視点の色で教示されるけど。
それをターゲットの色で教示するImmediate条件をつかって、淡蒼球外節から記録し。
ついでにNoGo試行もいれて、発火が上がる細胞と下がる細胞の機能を考察されてる。
いわれてみれば、antiのcueをターゲット自体にするってのは初めてみるかも。
素直に興味深い。
結果として、発火の上がるタイプは、やはりサッケードの駆動に関与してるっぽいけど。
下がるタイプの細胞は、ImmediateにするとantiだけでなくNoGoでも同様に下がるので。
間接路経由で眼球運動の調節に関わってる?
データはきれいなのに、あたし自身が基底核に疎いもんで。
最後の概念図のところを早くも失念してしまった。
(数日前に聞いた内容なのにこのザマだよorz)
P1-2-105 課題実行に伴う左右手選択意図を判別する*それをターゲットの色で教示するImmediate条件をつかって、淡蒼球外節から記録し。
ついでにNoGo試行もいれて、発火が上がる細胞と下がる細胞の機能を考察されてる。
いわれてみれば、antiのcueをターゲット自体にするってのは初めてみるかも。
素直に興味深い。
結果として、発火の上がるタイプは、やはりサッケードの駆動に関与してるっぽいけど。
下がるタイプの細胞は、ImmediateにするとantiだけでなくNoGoでも同様に下がるので。
間接路経由で眼球運動の調節に関わってる?
データはきれいなのに、あたし自身が基底核に疎いもんで。
最後の概念図のところを早くも失念してしまった。
(数日前に聞いた内容なのにこのザマだよorz)
雨宮 薫, 大須 理英子
発表者に出会わず、説明聞けなかった。
EEGの信号から運動肢をデコードしてるみたいだったんだけど。
なんでいまさら、そんなことをしようとしたのだろうか…。
P1-2-180 空腹と餌の選択:満足が引き起こす選択行動の乱雑さの増加EEGの信号から運動肢をデコードしてるみたいだったんだけど。
なんでいまさら、そんなことをしようとしたのだろうか…。
山田 洋
マーモセットをつかって、エサの選好性が空腹によって影響をうけるか検討。
結果としては、多くの個体で空腹時ほど一貫した選好傾向をみせ。
空腹じゃなければ、好みじゃないエサも結構選んだりする、という。
課題としては、5種類のエサの組み合わせで、2肢選択やらせて。
各ペアの選択率をもとに、選好を調べてるだけ。
解析はMDSとか推移律の崩れのカウントとか。
わたしも以前試した解析手順でした。
やっぱそういう方法しかないのかな。
三つ巴数えるやりかたは、かなり感度悪いので。
できれば選択率の量的関係を加味できる指標があるといいんだけど。
P1-2-183 海馬と扁桃体間のガンマオシレーションの同期は報酬予測による行動調節に寄与する*結果としては、多くの個体で空腹時ほど一貫した選好傾向をみせ。
空腹じゃなければ、好みじゃないエサも結構選んだりする、という。
「腹へってるときは、ともかくなんでもいいやって食う」
という予想とは、ある意味逆かな?課題としては、5種類のエサの組み合わせで、2肢選択やらせて。
各ペアの選択率をもとに、選好を調べてるだけ。
解析はMDSとか推移律の崩れのカウントとか。
わたしも以前試した解析手順でした。
やっぱそういう方法しかないのかな。
三つ巴数えるやりかたは、かなり感度悪いので。
できれば選択率の量的関係を加味できる指標があるといいんだけど。
寺田 慧, 高橋 晋, 櫻井 芳雄
正答時の報酬確率の高低をトーンで知らせつつ、ラットにノーズポーク課題をやらせ。
その間の海馬と扁桃体のLFPを調べた、と。
結果、報酬確率が高いときだけ、海馬-扁桃体のコヒーレンスがRTと相関した。
けっこう面白そうと思ったんだけど、混んでてちゃんと聞けずじまい。
P1-2-234 マウスの連続的レバー操作と運動野ニューロン活動との相互情報量推定その間の海馬と扁桃体のLFPを調べた、と。
結果、報酬確率が高いときだけ、海馬-扁桃体のコヒーレンスがRTと相関した。
けっこう面白そうと思ったんだけど、混んでてちゃんと聞けずじまい。
田中 康裕, 正水 芳人, 松崎 政紀
情報量解析はつねづねやりたいとおもってるので。
そのきっかけにと聞きにいったものの。
メインは
これはいまのとこ、あたしには関係ないな…。
そのきっかけにと聞きにいったものの。
メインは
・連続値同士の情報量を計算するにはCopulaのエントロピーを求めるが
・そのためにはデータのbinningが必要で、結果がbinサイズになるので
・AICによって適切なbinサイズを決めてみました
という内容だった。・そのためにはデータのbinningが必要で、結果がbinサイズになるので
・AICによって適切なbinサイズを決めてみました
これはいまのとこ、あたしには関係ないな…。
2日目(2013.06.21)
P2-1-126 サル下側頭皮質における倒立顔とサッチャー顔の集団コーディング松本 有央, 菅生-宮本 康子, 河野 憲二
顔全体が逆さだと、目・口がフリップしてても違和感が少ないサッチャー錯視。
顔のような複雑な形態情報は、下側頭葉で行なわれているので。
サッチャー顔を含む顔刺激をみせつつITニューロンを記録するという。
それをサルでやるという発想がすげーな。
結果として、ITのニューロンは顔をコードしてて。
正立像だと、目・口のフリップを検出するんだけど。
倒立したサッチャー顔では、非サッチャー顔との弁別性が下がる。
つまりニューロンレベルで、個体の知覚と似たような性質をもっている。
しかもヒトはヒトの顔、サルはサルの顔でしか目・口のフリップに違和感を感じないが。
サルITニューロンもおなじく、ヒトの顔ではフリップにsensitiveではない。
面白いねこれは。
P2-1-132 情報量最大化原理によるV2の選択性の説明顔のような複雑な形態情報は、下側頭葉で行なわれているので。
サッチャー顔を含む顔刺激をみせつつITニューロンを記録するという。
それをサルでやるという発想がすげーな。
結果として、ITのニューロンは顔をコードしてて。
正立像だと、目・口のフリップを検出するんだけど。
倒立したサッチャー顔では、非サッチャー顔との弁別性が下がる。
つまりニューロンレベルで、個体の知覚と似たような性質をもっている。
しかもヒトはヒトの顔、サルはサルの顔でしか目・口のフリップに違和感を感じないが。
サルITニューロンもおなじく、ヒトの顔ではフリップにsensitiveではない。
面白いねこれは。
藤原 謙三郎, 田中 琢真, 中村 清彦
3層からなる人工ニューラルネットワークに、自然画像を入力としてあたえ。
各層の出力のエントロピーが最大になるよう、結合強度を学習させてやる。
すると、
個人的にこういう研究好きなので、面白い。
が、肝心のV2ライクな受容野のデータが少ないし。
そもそもあんまりきれいな受容野構造にはみえない。
その応答特性は、1層のGabor方位の重ね書きでは綺麗に描けないような。
面の処理に関わる受容野を定量する方法が欲しいところ。
P2-2-178 サル下側頭葉皮質と前頭前野からの皮質脳波を用いて、想起された視覚記憶をデコーディングする各層の出力のエントロピーが最大になるよう、結合強度を学習させてやる。
すると、
・1層のユニットはほとんどがGabor様の受容野(simple cellに類似)を
・2層のユニットはより広い方位選択性の受容野(complex cellに類似)を
・3層のユニットはエッジを検出するかのような複雑な受容野(V2 cellに類似)をもった
という研究。・2層のユニットはより広い方位選択性の受容野(complex cellに類似)を
・3層のユニットはエッジを検出するかのような複雑な受容野(V2 cellに類似)をもった
個人的にこういう研究好きなので、面白い。
が、肝心のV2ライクな受容野のデータが少ないし。
そもそもあんまりきれいな受容野構造にはみえない。
「V2にはテクスチャーでつくられたエッジなどに応答する細胞がいる」
ってハナシをされてたけど…。その応答特性は、1層のGabor方位の重ね書きでは綺麗に描けないような。
面の処理に関わる受容野を定量する方法が欲しいところ。
谷川 久, 長谷川 功
サル下側頭葉と前頭前野にECoG電極を埋め込み。
対連合課題中の信号から、視覚情報の想起過程をデコードしよう、という研究。
実験のつくりとしては非常にかたいんだけど。
各時間binで訓練したデコーダで、そのbinの活動をデコードしてるのはなぜだろう。
それだと、時間毎でデコーダは独立になっちゃうし。
pair-recallみたいな細胞とのアナロジーで、
まあワーキングメモリ課題を課した場合との比較もしてあって。
記憶課題で訓練したデコーダは、対連合課題の信号をデコード不可ってことだったので。
いちおう「みせた刺激をデコードしてるだけではない」ってコトか。
P2-2-179 長期記憶想起におけるサル内側側頭皮質脳波の周波数間結合対連合課題中の信号から、視覚情報の想起過程をデコードしよう、という研究。
実験のつくりとしては非常にかたいんだけど。
各時間binで訓練したデコーダで、そのbinの活動をデコードしてるのはなぜだろう。
それだと、時間毎でデコーダは独立になっちゃうし。
pair-recallみたいな細胞とのアナロジーで、
・paired associatesをみせてるときの活動でデコーダを訓練して
・そのデコーダを、遅延期間の信号に適用する
みたいなやりかたではダメなのかしら。・そのデコーダを、遅延期間の信号に適用する
まあワーキングメモリ課題を課した場合との比較もしてあって。
記憶課題で訓練したデコーダは、対連合課題の信号をデコード不可ってことだったので。
いちおう「みせた刺激をデコードしてるだけではない」ってコトか。
安達 賢, 長谷川 功, 中原 潔
上記とおなじグループのポスターで。
こちらはECoG電極が下側頭葉だけ。
で、LFP自体でも刺激選択性とかをみてるんですが。
面白いのは、遅延期中にシータ波の位相とガンマ振幅のカップリングがあった。
つまり一部のチャンネルで、シータがある位相のときにガンマ振幅が増えていた、と。
シータ波というと、とりあえず海馬が思い浮かびますが。
いまのところ解析は同一チャンネル内でしかしていないそうで、ちょい残念。
せっかくECoGで広くとってるんだし。
ITのガンマにいちばん強く影響するシータ源さがすとか、いろいろできそう。
前頭前野と低次視覚野のデータもあれば、なお面白い。
P2-2-189 マカクザルにおける統計的単語分割に関する神経相関の特定こちらはECoG電極が下側頭葉だけ。
で、LFP自体でも刺激選択性とかをみてるんですが。
面白いのは、遅延期中にシータ波の位相とガンマ振幅のカップリングがあった。
つまり一部のチャンネルで、シータがある位相のときにガンマ振幅が増えていた、と。
シータ波というと、とりあえず海馬が思い浮かびますが。
いまのところ解析は同一チャンネル内でしかしていないそうで、ちょい残念。
せっかくECoGで広くとってるんだし。
ITのガンマにいちばん強く影響するシータ源さがすとか、いろいろできそう。
前頭前野と低次視覚野のデータもあれば、なお面白い。
田村 潤, 池田 和司, 藤井 直敬
いくつかの高さのトーンを「音素」とみなし。
その3つを組み合わせた3音の系列を「単語」とみなす。
そうやって人工的につくった数種の単語を、区切りなく延々とサルに聞かせ。
ECoG信号から、単語間の区切りの認識に関する処理過程を調べるという実験。
うーん。
まあ発想は面白いとおもうんだけど。
これって結局は、トーン系列の頻度の違いによるオドボールなので。
単語処理云々という題目で言語機能のセクションに出すのは、この時点ではいかがなものか。
そういう思想があっての実験だということは理解するけれど。
P2-2-234 運動野のマルチニューロン活動に対する経頭蓋直流電気刺激の影響その3つを組み合わせた3音の系列を「単語」とみなす。
そうやって人工的につくった数種の単語を、区切りなく延々とサルに聞かせ。
ECoG信号から、単語間の区切りの認識に関する処理過程を調べるという実験。
うーん。
まあ発想は面白いとおもうんだけど。
これって結局は、トーン系列の頻度の違いによるオドボールなので。
単語処理云々という題目で言語機能のセクションに出すのは、この時点ではいかがなものか。
そういう思想があっての実験だということは理解するけれど。
田中 智子, 本田 学
ラットにtDCSをかけてやり、そのときの皮質ニューロンの活動をみた研究。
地味だけど必要な仕事ですね。
ただ、データが少ないせいもあってか、結果はいまのところ議論の余地ありで。
一部の細胞種のみ、tDCSで発火頻度の変化が起きたが。
活動増加とも減少ともつかない微妙なケースのほうが多いみたい。
(アブストにはMUAのハナシしか記載されてないから、詳しく書くわけにいかん。)
とりあえずもっとデータ増やしてどうなるか、ってカンジだろうか。
地味だけど必要な仕事ですね。
ただ、データが少ないせいもあってか、結果はいまのところ議論の余地ありで。
一部の細胞種のみ、tDCSで発火頻度の変化が起きたが。
活動増加とも減少ともつかない微妙なケースのほうが多いみたい。
(アブストにはMUAのハナシしか記載されてないから、詳しく書くわけにいかん。)
とりあえずもっとデータ増やしてどうなるか、ってカンジだろうか。
3日目(2013.06.22)
P3-1-134 補足眼野で生成される予測誤差は探索と知識利用の切り替えを引き起こす*川口 典彦, 坂本 一寛, 虫明 元
4つのターゲットのうち2つが正解となる試行錯誤な探索課題で。
試行錯誤中の「はずれ」や、分かりかけてる「あたり」に応答するニューロンがいる。
とってるのは補足眼野。
小川先生のラボの「異なるエラー表現」のはなしに似てて面白いし。
できれば詳しく聞きたかったが。
自分のポスターがあったため、ちょろっとだけしか聞けず。
P3-1-136 サリエンシー・マップに基づいた視覚補填を目指して*試行錯誤中の「はずれ」や、分かりかけてる「あたり」に応答するニューロンがいる。
とってるのは補足眼野。
小川先生のラボの「異なるエラー表現」のはなしに似てて面白いし。
できれば詳しく聞きたかったが。
自分のポスターがあったため、ちょろっとだけしか聞けず。
吉田 正俊, Richard Veale
以前もどこかで耳にした気はするんだけど。
上丘刺激で注意を駆動することで、視覚障害の人工補綴につかえないか、という研究。
とはいえ、ホントにそういうことやったデータではなく。
とりあえずビデオ映像をもとに「現在の視野のsaliencyマップ」をリアルタイムで計算し。
それに対応する上丘領域を刺激して、注意をその位置へ誘導できるかのシミュレーション。
けっこう人だかりになってたみたいなのだが。
これも聞いてる余裕がなくて、ちゃんと聞けずじまい。
P3-2-51 確率情報及び意思決定の履歴が知覚的意思決定に作用する神経機構上丘刺激で注意を駆動することで、視覚障害の人工補綴につかえないか、という研究。
とはいえ、ホントにそういうことやったデータではなく。
とりあえずビデオ映像をもとに「現在の視野のsaliencyマップ」をリアルタイムで計算し。
それに対応する上丘領域を刺激して、注意をその位置へ誘導できるかのシミュレーション。
けっこう人だかりになってたみたいなのだが。
これも聞いてる余裕がなくて、ちゃんと聞けずじまい。
金子 宜之, 梅田 和昌, 坂井 克之
正弦縞が下に動いたかどうかを報告させる知覚的意思決定課題を用い。
確率知識および先行試行での選択が、現在の判断に与える影響を調べる。
ついでにfMRIで責任領域を割り出す、という仕事。
とりあえず信号検出理論をつかった行動解析で、確率知識は判断基準(C)をバイアスし。
一方で先行する選択は、弁別能(d')に影響する。
そのうえでfMRI信号と被験者ごとの行動指標の相関を取ると。
確率知識と先行試行の影響それぞれについて、異なる領域の信号強度と相関が得られた。
(抄録には左STSのことしか書かれてないので、詳細は省略。)
まあ、最後の責任領域のハナシについては、どこまで信憑性あるかわからないが。
少なくとも行動のデータは、自分の研究に関連する内容なので。
なんでこういう効果になるのか、考えておく必要ありだわ。
P3-2-52 外側手綱核と前部帯状皮質における学習シグナルの表現確率知識および先行試行での選択が、現在の判断に与える影響を調べる。
ついでにfMRIで責任領域を割り出す、という仕事。
とりあえず信号検出理論をつかった行動解析で、確率知識は判断基準(C)をバイアスし。
一方で先行する選択は、弁別能(d')に影響する。
そのうえでfMRI信号と被験者ごとの行動指標の相関を取ると。
確率知識と先行試行の影響それぞれについて、異なる領域の信号強度と相関が得られた。
(抄録には左STSのことしか書かれてないので、詳細は省略。)
まあ、最後の責任領域のハナシについては、どこまで信憑性あるかわからないが。
少なくとも行動のデータは、自分の研究に関連する内容なので。
なんでこういう効果になるのか、考えておく必要ありだわ。
川合 隆嗣, 高田 昌彦, 松本 正幸
間欠強化なスケジュールで逆転学習課題をサルに行なわせ。
その間の外側手綱核と前帯状回の単一ニューロン活動記録。
結果として、手綱核は基本的に負の強化子をそのまま表現してるだけの応答が多く。
いっぽう前帯状回は、逆転後の無報酬経験の回数増加により、活動が漸増したりする。
つまり後者のほうが、学習に関わってるっぽい、と。
ところで、ふと気になって
やっぱ手綱核狙うときは、かなり静脈洞のキワキワに刺さなきゃならないから。
最初に刺すときはどこまでがセーフかわからないから、かなり怖いとのことでした。
やっぱそうなのかー。
P3-2-53 行動決定課題遂行における報酬価値情報処理に関連したアカゲザル眼窩前頭皮質のニューロン活動その間の外側手綱核と前帯状回の単一ニューロン活動記録。
結果として、手綱核は基本的に負の強化子をそのまま表現してるだけの応答が多く。
いっぽう前帯状回は、逆転後の無報酬経験の回数増加により、活動が漸増したりする。
つまり後者のほうが、学習に関わってるっぽい、と。
ところで、ふと気になって
「うっかり静脈洞刺したりとか、ないんですか?」
とか聞いてみたら。やっぱ手綱核狙うときは、かなり静脈洞のキワキワに刺さなきゃならないから。
最初に刺すときはどこまでがセーフかわからないから、かなり怖いとのことでした。
やっぱそうなのかー。
瀬戸川 剛, 設楽 宗孝
n回の試行を立て続けに終えると、m滴ジュースがもらえる課題をサルに訓練し。
その後、nとmをそれぞれ1/2/4の組み合わせで振って、サル自身に選ばせる。
たとえば、
で、その選択をしてるときの前頭眼窩野の活動を記録したという研究。
OFCだから、当然報酬予測に関わる応答が出るわけですが。
とくに面白いタイプの応答としては、
いずれもニューロンの数としては少なかったので。
もっとデータ集まったときに、どういう割合になるかとかは、いまのとこ未知数。
それより解析対象とした期間が気になりました。
今回は選択肢を系列呈示してる期間と、そのあとの選択期の活動を解析してたが。
選択肢によっては、それが出た瞬間に選択が決定するようなヤツもあるはずなんで。
(明らかに得または損な選択肢。)
そういうのが出た直後の遅延期とか調べたほうが、面白いんじゃなかろうか。
P3-2-57 自由選択課題時における線条体の階層的行動表現その後、nとmをそれぞれ1/2/4の組み合わせで振って、サル自身に選ばせる。
たとえば、
「1試行で1滴と、2試行やって4滴、どっちがいい?」
みたいな。で、その選択をしてるときの前頭眼窩野の活動を記録したという研究。
OFCだから、当然報酬予測に関わる応答が出るわけですが。
とくに面白いタイプの応答としては、
・ひとつめとふたつめの選択肢の価値の差に相関するニューロン
・選択肢間の価値の差が小さいほど応答するニューロン(コンフリクトを表現)
・逆に差が大きいほど応答するニューロン
などなど。・選択肢間の価値の差が小さいほど応答するニューロン(コンフリクトを表現)
・逆に差が大きいほど応答するニューロン
いずれもニューロンの数としては少なかったので。
もっとデータ集まったときに、どういう割合になるかとかは、いまのとこ未知数。
それより解析対象とした期間が気になりました。
今回は選択肢を系列呈示してる期間と、そのあとの選択期の活動を解析してたが。
選択肢によっては、それが出た瞬間に選択が決定するようなヤツもあるはずなんで。
(明らかに得または損な選択肢。)
そういうのが出た直後の遅延期とか調べたほうが、面白いんじゃなかろうか。
伊藤 真, 銅谷 賢治
ラットに並列間欠強化スケジュールをやらせ。
その間の線条体の活動を記録。
いつもはここから、反応方向や報酬情報、価値関数を表現する活動を報告されてるけど。
今回は、ポピュレーションの活動から、試行内の時間をデコードするという。
ちょっと違う切り口。
cross validationのため、データを分割し。
そのうえで活動は100msのbinで切る。
結果としては、線条体ニューロンの活動パターンから、試行内時間をデコードできた、と。
とくに背内側領域が、デコーディング精度高め。
とはいえ、そもそも特定のイベントで応答する課題関連ニューロンがいるわけだから。
それらの活動を参照すれば、いま試行内のどこにいるか、そりゃわかるだろう。
なので、この解析で何がわかったのか、いまいちあたしには理解できず。
まあ、そもそもこういう解析をするモチベーションとして、
その気持ちはわかるけどね。
P3-2-60 基底核および小脳における時間情報の神経表現その間の線条体の活動を記録。
いつもはここから、反応方向や報酬情報、価値関数を表現する活動を報告されてるけど。
今回は、ポピュレーションの活動から、試行内の時間をデコードするという。
ちょっと違う切り口。
cross validationのため、データを分割し。
そのうえで活動は100msのbinで切る。
結果としては、線条体ニューロンの活動パターンから、試行内時間をデコードできた、と。
とくに背内側領域が、デコーディング精度高め。
とはいえ、そもそも特定のイベントで応答する課題関連ニューロンがいるわけだから。
それらの活動を参照すれば、いま試行内のどこにいるか、そりゃわかるだろう。
なので、この解析で何がわかったのか、いまいちあたしには理解できず。
まあ、そもそもこういう解析をするモチベーションとして、
「ニューロンみてると、試行内の中途半端なタイミングで活動するヤツがいる」
「そういう細胞って、いったいなにやってんのかなって思った」
っておっしゃってたので。「そういう細胞って、いったいなにやってんのかなって思った」
その気持ちはわかるけどね。
國松 淳, 田中 真樹
いくつかの異なる長さ注視をつづけ、その後self-timingでサッケードをさせる。
その課題中での、基底核と小脳歯状核の応答の比較。
基底核の細胞は、いままでどおり、サッケードの開始目指してだんだん活動をあげるので。
遅延が長いほど、ゆっくりとしたスロープでbuild-upする。
一方小脳は、だいたいがサッケード直前になって急激に活動をあげるので。
スロープの角度は、条件間であまりかわらない、と。
それはまあ、いいとおもうんだけど。
この結果から、
時間表現っていうより、単に運動の駆動じゃね?
小脳だからといってintermediate timeの処理と結びつける必然性はないのでは。
P3-2-63 リスクを伴う意思決定における島皮質前部の神経表現*その課題中での、基底核と小脳歯状核の応答の比較。
基底核の細胞は、いままでどおり、サッケードの開始目指してだんだん活動をあげるので。
遅延が長いほど、ゆっくりとしたスロープでbuild-upする。
一方小脳は、だいたいがサッケード直前になって急激に活動をあげるので。
スロープの角度は、条件間であまりかわらない、と。
それはまあ、いいとおもうんだけど。
この結果から、
「小脳はより精密な時間の表現に関与」
っていう解釈になるのは、どうなのか。時間表現っていうより、単に運動の駆動じゃね?
小脳だからといってintermediate timeの処理と結びつける必然性はないのでは。
石井 宏憲, 筒井 健一郎, 飯島 敏夫
「確実な小報酬」と「確率的な大報酬」の選択課題をラットに行なわせ。
前島皮質(など)から記録したり、inactivationしたり。
単純なギャンブリング課題だけど、けっこう面白いと思ったんだが。
あまりシステマティックな解釈は、いまのとこできないカンジなのだろうか。
細胞をタイプ分類してたが、それらをどう解釈するのか、いまひとつ分からずじまい。
P3-2-64 課題条件に応じた行動選択時のラットの前頭前野と背側線条体の役割前島皮質(など)から記録したり、inactivationしたり。
単純なギャンブリング課題だけど、けっこう面白いと思ったんだが。
あまりシステマティックな解釈は、いまのとこできないカンジなのだろうか。
細胞をタイプ分類してたが、それらをどう解釈するのか、いまひとつ分からずじまい。
船水 章大, 銅谷 賢治, 高橋 宏知
これまでも何度か神経科学大会で、似たデータの発表聞いてる気がするけど。
どこが新しくなったのだろうか。
prelimbicの活動のデータとか出てるのは、初でしたっけ?
P3-2-67 複数の特徴次元に由来する視覚的顕著性信号の計算様式*どこが新しくなったのだろうか。
prelimbicの活動のデータとか出てるのは、初でしたっけ?
大渕 藍, 小川 正
小川先生のラボのプロジェクトのなかでは、唯一よく知らないヤツだと思うので。
説明してもらいたかったんだけど。
ついぞ発表者が立ってるのをみかけなかった。
P3-2-69 行動選択におけるラット前頭前野の多細胞集団活動の挙動説明してもらいたかったんだけど。
ついぞ発表者が立ってるのをみかけなかった。
半田 高史, 礒村 宜和, 深井 朋樹
高音なら左、低音なら右のリッキングを頭固定のラットに訓練しておき。
中間音を聞かせたとき、動物の行動とmPFCの活動を記録。
流行りのstate spaceとか発火パターンの距離を使った解析が多く、面白かった。
まあその解析で、どれほどユニークなことが分かるかは、ちょっと不明ながら。
こういうでかい集団データをどう扱うか、勉強になったというか。
そこで線形判別つかえばいいのかー、みたいな。
P3-2-74 報酬に基づく行動のバイアスは視床CM核―線条体投射によって最適化される*中間音を聞かせたとき、動物の行動とmPFCの活動を記録。
流行りのstate spaceとか発火パターンの距離を使った解析が多く、面白かった。
まあその解析で、どれほどユニークなことが分かるかは、ちょっと不明ながら。
こういうでかい集団データをどう扱うか、勉強になったというか。
そこで線形判別つかえばいいのかー、みたいな。
山中 航, 木村 實
ふたつの反応対象に、それぞれ異なる報酬確率を連合しておき。
あくまで反応は、刺激によって示したほうの選択肢にさせる。
それでもサルとしては、高報酬確率の運動を「よりしたい」というバイアスを抱くので。
それが視床CM核の応答にどう影響するか、みたいなハナシ?
課題は単純なのに、データはかなり面白そうだったんですが。
いかんせん立ちどおしで、腰が限界に近くて。
詳しく説明聞けなかったorz
P3-2-242 一次運動野に導入した光感受性膜タンパク質による、行動中のマカクザル前肢筋活動の光遺伝学的抑制あくまで反応は、刺激によって示したほうの選択肢にさせる。
それでもサルとしては、高報酬確率の運動を「よりしたい」というバイアスを抱くので。
それが視床CM核の応答にどう影響するか、みたいなハナシ?
課題は単純なのに、データはかなり面白そうだったんですが。
いかんせん立ちどおしで、腰が限界に近くて。
詳しく説明聞けなかったorz
木下 正治, 伊佐 正
サルの運動野にAAVでハロを発現させてやり。
光刺激で腕の筋電に影響出たという内容。
腕全体の運動にはハッキリとした効果は出ておらず。
筋電にしても、何千試行と加算して、やっとキレイにみえてくるレベルですが。
とはいえサルの運動野では、光遺伝的制御の影響ってまだ報告されてない気がするので。
たぶん近いうちに、どっかに発表されるだろう。
ただ、やっぱこんだけ効果が小さいと。
それをどう使うかってトコがね。
サルでの光刺激なんかは、やっぱいまだ発展途上ってことか。
光刺激で腕の筋電に影響出たという内容。
腕全体の運動にはハッキリとした効果は出ておらず。
筋電にしても、何千試行と加算して、やっとキレイにみえてくるレベルですが。
とはいえサルの運動野では、光遺伝的制御の影響ってまだ報告されてない気がするので。
たぶん近いうちに、どっかに発表されるだろう。
ただ、やっぱこんだけ効果が小さいと。
それをどう使うかってトコがね。
サルでの光刺激なんかは、やっぱいまだ発展途上ってことか。
感想
えー、感想。すごく……腰が痛いです……。
おもに3日目のハードスケジュールのせいで。
というのも、最初に書いたとおり、今年はポスターが午前・午後の貼り替え制で。
いつものように、空き時間にゆっくり見てまわるというのがしづらく。
短い時間を惜しむように、ガツガツみてまわらなければなりませんでした。
とくにあたしは、3日目の午前が自分のポスターで。
しかもその午後には、みたいポスターが集中していたので。
終わってみれば、3日目はポスター会場以外いってないという。
もちろんその間、だらだらまわってたわけではなく。
チェックしたいとこをピンポイントでまわったのに、ポスターだけで一日終了。
まあ考えてみれば、貼り替え制ってことは、単純に一日のポスター枚数が例年の倍なわけで。
そりゃ腰もやられるわ。
しかしそうして、ある意味「詰め込んだ」おかげなのか。
今年はポスター会場の活気が、例年よりも熱かった印象でした。
コアタイム以外の時間も、わりとひとだかりがあったカンジ。
なので、閑散としたポスター会場をだらだらみるのが好きな、あたしみたいのは別としても。
一般論として貼り替え制は、ポスター討論の促進という効果を生んだ気がします。
ただ、それで万事ハッピーだったかというと、そんなことはないです。
まず理解不能なのが、午前と午後でポスター掲示の長さがぜんぜん違うこと。
午前は8:45に貼って12:35に剥がすから、持ち時間は約4時間。
しかし午後は14:00に貼って19:00まで置いとけるから、持ち時間は5時間。
なんで1時間も違うのよ。
しかも冊子のタイムテーブルでは、じつに子供だましな小細工がされていて。
午後の最後の1時間は「ポスター閲覧」の色塗りがされておらず。
一見すると午前と午後の長さが同じにみえるんですよ。
ざけてんのか。
あともうひとつ、これもまったく理解に苦しむのですが。
例年プレナリーの時間帯は、他のシンポやオーラルはぶつけられてなくて。
プレナリー聞くか、ポスター会場ぶらつくかの二択なカンジなんですが。
今年はなんと、プレナリーの1時間、ポスターまでもが空白の時間にされていて。
その間はポスターを貼っていてはいけないという意味不明さ。
(午前のポスター撤去と午後の貼り付けの時間は、ちゃんと別にとってあるのに。)
そのためプレナリーを聞かないと、例年のようにポスターをみてまわることもできず。
ただただ無駄に1時間を、他になにもやってない会場でつぶすことになる。
あたしは一日もプレナリー聞かなかったので。
毎日、昼の1時間だけぽっかりヒマという、理不尽な思いをしました。
(聞きたくもないプレナリーに無理に入っても、寝ちゃうだけだしね。)
ということで。
ポスターが半日ごと入替制だったのは、善し悪し双方あったかもしれませんが。
日程全体としては、ハッキリいって腐れ仕様。
今年のタイムテーブルつくった輩は、もう[自主規制]とおもいます。
ま、今年は三学会合同大会ゆえの、って部分もあるはずなので。
来年はもうちょいまともな運営になってくれることを期待します。