@kanri_ninjin
@kanri_ninjin


 イクナイ! 581
イクナイ! 581







第35回 日本神経科学大会(2012)の記録
2012年の神経科学学会に参加した際の記録です。見聞きしたことを忘れないための、至極個人的なメモです。
理解不足や間違え等もあるかとおもいますので、悪しからず。
発表者が複数いる場合、ファーストとラスト、あるいはラストに準ずるひと2~3人だけ表記します。
(みやすさの優先のため。)
アスタリスクつきのポスターは、自分でちょろっと読んだだけのヤツ。
アスタリスクなしは、ちゃんと発表者に説明してもらったヤツです。
基本情報
会場は名古屋国際会議場。2009年と同じ場所ですね。
大会長は名古屋大の貝淵弘三先生。
今大会より、プログラム冊子の送付すらなくなり。
事前に送られてくるのは、白紙のネームカードとプログラム引換券のみ。
大会会場にて、その引換券と交換で冊子をもらえるシステムになりました。
まあ合理的なのはわかるけど…。
事前に冊子にいろいろ書き込んどくとかできなくて不便。
(´д`)
それから、今年から大会が本格的な4日間スケジュールになりました。
まあ、いちおう去年も4日間だったとはいえ。
アレは初日が夕方の講演のみっていう、意味不明構成だったけど。
今年は、1日目の朝イチから4日目の18時までのミッチリ。
ま、ホントに4日間キツキツだったかというと、そうでもなくて。
いちおうシンポジウムなんかは全日にばらけているんだけど。
口頭発表が多かったり、中日の3日目はポスター数半分だったり。
とくにわれわれ認知屋には、2日目と3日目がえらい不作で。
必ずしも「充実の4日間」ではなかったのですが。
来年からもずっと4日間安定でいくのだろうか…。
しかも来年は、3学会合同大会の年だしね。
スケジュール
4日間のあたしの動き。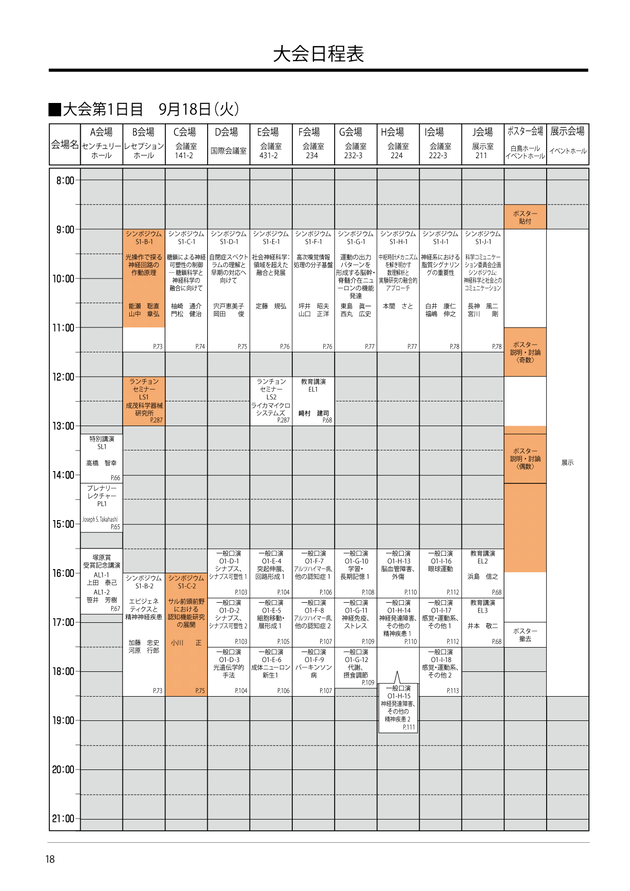



トーク
1日目(2012.09.18)
S1-B-1 光操作で探る神経回路の作動原理能瀬 聡直, 山中 章弘
小宮山さんの発表に期待していったんだけど。
おもに嗅球のイメージングのハナシだった。
嗅覚をつかった行動課題とかでもなく。
「感覚生理学」的な。
うーむ。
しょうがないので、4人目の山中章弘先生まで聞いて、出てきてしまいました。
なぜか初日だけ、シンポが11:30までやってて。
午前のポスターコアタイムにかぶってたもんで。
LS1 脳の発生と進化の原理と多様性おもに嗅球のイメージングのハナシだった。
嗅覚をつかった行動課題とかでもなく。
「感覚生理学」的な。
うーむ。
しょうがないので、4人目の山中章弘先生まで聞いて、出てきてしまいました。
なぜか初日だけ、シンポが11:30までやってて。
午前のポスターコアタイムにかぶってたもんで。
岡野 栄之
ナリシゲのランチョンセミナー。
前大会長の大隅先生がチェアで。
講演本体は、慶応の岡野先生。
マーモセットの遺伝子改変技術など、たいへん興味深い内容だった。
いまのところ、マーモの遺伝学は病理学のほうへ進んでいるカンジで。
それは間違いではないとおもうんだけど。
方法論が一般的になれば、認知神経科学にもいろんな貢献があるとおもう。
神経疾患の中間表現型のなかには、いろんな認知機能が含まれるからね。
それにしても、岡野先生のトークはいつもおもしろいなぁ。
いろんな実験結果をつぎつぎと明解に説明されるので。
S1-C-2 サル前頭前野における認知機能研究の展開前大会長の大隅先生がチェアで。
講演本体は、慶応の岡野先生。
マーモセットの遺伝子改変技術など、たいへん興味深い内容だった。
いまのところ、マーモの遺伝学は病理学のほうへ進んでいるカンジで。
それは間違いではないとおもうんだけど。
方法論が一般的になれば、認知神経科学にもいろんな貢献があるとおもう。
神経疾患の中間表現型のなかには、いろんな認知機能が含まれるからね。
それにしても、岡野先生のトークはいつもおもしろいなぁ。
いろんな実験結果をつぎつぎと明解に説明されるので。
「すごく勉強になるハナシを聞いた」
という気にさせられる。小川 正, Jonathan D. Wallis, 船橋 新太郎
今年のシンポのなかで、おそらくもっともあたしに関係あるセッション。
…だったんだけど、なんというか結構ビミョーだった。
小川先生と辻本先生のトークは、内容としては明解でよかったんですが。
いかんせん、いままで何度も聞いたことのあるデータだったから。
とくにあらたな感想もなく。
星先生とJonathan Wallisは、なかなかおもしろかったけど…。
最後の船橋先生は、対連合課題の説明とかまったくしてないうちから
けっこうな数のひとが席を立ってて、いたたまれないカンジだった。
やっぱ認知屋は、
それ抜きで課題とかニューロン活動とか説明されても、何がいいたいのかサッパリ。
自分は気をつけようと、改めておもった次第です。
…だったんだけど、なんというか結構ビミョーだった。
小川先生と辻本先生のトークは、内容としては明解でよかったんですが。
いかんせん、いままで何度も聞いたことのあるデータだったから。
とくにあらたな感想もなく。
星先生とJonathan Wallisは、なかなかおもしろかったけど…。
最後の船橋先生は、対連合課題の説明とかまったくしてないうちから
「paired associateが呈示されたときのニューロンの発火が…」
とかいい始めちゃうし。けっこうな数のひとが席を立ってて、いたたまれないカンジだった。
やっぱ認知屋は、
自分がどういう認知機能に興味があって、研究の目的はなんなのか
をとにかくハッキリさせないと。それ抜きで課題とかニューロン活動とか説明されても、何がいいたいのかサッパリ。
自分は気をつけようと、改めておもった次第です。
2日目(2012.09.19)
EL4 教育講演4高田 昌彦, 下郡 智美
今年からはじまった、日本語による教育講演企画。
なかなかどうして面白そうなのが多かったのですが。
いかんせん、別のシンポとぶつかってしまうものも多く。
このEL4は昼食時間にやってたので、ランチョンは捨てて聞きにいきました。
高田先生は、順向性・逆向性のウイルスベクタをもちいた遺伝子導入のハナシ。
投射先と投射元からウイルスをいれて、特定の回路にだけ遺伝子を発現させる。
2人目の下郡先生は、いつものin uteroのハナシ。
どちらも、現在の認知神経科学において欠かせない技術なワケですが。
現時点でのあたしの立場としては、前者のほうが現実味をもって重要かなぁ。
ある程度ラインが確立してしまえば、in uteroする必要もなくなっちゃうしね。
致死性変異なら別だけど、認知屋はそういう遺伝子は対象にしないし。
O2-I-35 感覚・運動系、その他 4なかなかどうして面白そうなのが多かったのですが。
いかんせん、別のシンポとぶつかってしまうものも多く。
このEL4は昼食時間にやってたので、ランチョンは捨てて聞きにいきました。
高田先生は、順向性・逆向性のウイルスベクタをもちいた遺伝子導入のハナシ。
投射先と投射元からウイルスをいれて、特定の回路にだけ遺伝子を発現させる。
2人目の下郡先生は、いつものin uteroのハナシ。
どちらも、現在の認知神経科学において欠かせない技術なワケですが。
現時点でのあたしの立場としては、前者のほうが現実味をもって重要かなぁ。
ある程度ラインが確立してしまえば、in uteroする必要もなくなっちゃうしね。
致死性変異なら別だけど、認知屋はそういう遺伝子は対象にしないし。
永雄 総一
初日に電車の駅で、磯村研の齊木さんにバッタリお会いしまして。
そこで、同研究室の木村さんがオーラルで出してるとお聞きしたので。
聞きにいってきました。
木村さんはセッションの2人目でしたが。
1人目の五十嵐さんも、以前ポスターを説明していただいたことがあったので。
このお二方のトコだけ。
五十嵐さんは、前に聞いたのより少し解析が進んだ印象かな。
磯村先生のデータのLFP解析のハナシ。
細胞種および層ごとに、スパイクとLFPの位相ロックを調べておられるんだけど。
やっぱそれがどう運動に関係しているかは、ちとあたしには解釈つかないのよね…。
木村さんのほうは、おなじく頭固定ラットにレバー引きをさせるんだけど。
タイミングを外的に指示する条件と、ラットに自発的にさせる条件があって。
各条件間でのM1・M2細胞活動の比較。
まー失礼になってしまうかもしれませんが、みたカンジ
spike coincidenceをどうストーリーにつなげてくかがポイントになりそうな。
とくに内発性の運動企図って、オンセットがそろわないので。
(オフセットは運動でトリガすればそろうけど。)
運動準備と発現のシナリオにどうのってくるか、今後注目なカンジです。
そうそう、そういや会場に外山先生がいらっしゃって。
いつもの調子で厳しいツッコミをいれておられたのですが。
なんかご用事でもあったのか、木村さんに
恐ろしすぎるだろアレ…
S2-B-2 神経生理学のための新しい方法:分子から装置までそこで、同研究室の木村さんがオーラルで出してるとお聞きしたので。
聞きにいってきました。
木村さんはセッションの2人目でしたが。
1人目の五十嵐さんも、以前ポスターを説明していただいたことがあったので。
このお二方のトコだけ。
五十嵐さんは、前に聞いたのより少し解析が進んだ印象かな。
磯村先生のデータのLFP解析のハナシ。
細胞種および層ごとに、スパイクとLFPの位相ロックを調べておられるんだけど。
やっぱそれがどう運動に関係しているかは、ちとあたしには解釈つかないのよね…。
木村さんのほうは、おなじく頭固定ラットにレバー引きをさせるんだけど。
タイミングを外的に指示する条件と、ラットに自発的にさせる条件があって。
各条件間でのM1・M2細胞活動の比較。
まー失礼になってしまうかもしれませんが、みたカンジ
「なんぞいろいろ出てきた」
って段階なのでしょうか。spike coincidenceをどうストーリーにつなげてくかがポイントになりそうな。
とくに内発性の運動企図って、オンセットがそろわないので。
(オフセットは運動でトリガすればそろうけど。)
運動準備と発現のシナリオにどうのってくるか、今後注目なカンジです。
そうそう、そういや会場に外山先生がいらっしゃって。
いつもの調子で厳しいツッコミをいれておられたのですが。
なんかご用事でもあったのか、木村さんに
「synchronizationとspike coincidenceの違いは!?」
って質問を投げ、そのまま轢き逃げのように会場を後にされたwwwww恐ろしすぎるだろアレ…
村山 正宜, 宮脇 敦史
前述のオーラルとかぶってたので、途中一部抜けましたが。
理研BSIの村山先生のシンポ。
個人的には、3人目のAlert Leeの発表が面白かった。
あたし、場所細胞のハナシはぜんぜん門外漢なんですが。
細胞内記録のデータから論理的に仮説を絞っていく過程が、すごくスマート。
ちなみに肝心の村山先生のトークは、ちょうど抜けてたので聞けませんでしたorz
理研BSIの村山先生のシンポ。
個人的には、3人目のAlert Leeの発表が面白かった。
あたし、場所細胞のハナシはぜんぜん門外漢なんですが。
細胞内記録のデータから論理的に仮説を絞っていく過程が、すごくスマート。
ちなみに肝心の村山先生のトークは、ちょうど抜けてたので聞けませんでしたorz
3日目(2012.09.20)
S3-J-1 古い脳に潜む新たな機能-故きを温ねて新しきを知る-山本 慎也, 橘 吉寿
なんか呼吸中枢のハナシとかはじまりそうなセッション名ですが。
どうやらそういうことではないようで。
皮質下構造や小脳を対象としてはいるものの、あくまで認知屋な内容です。
というか、3人目の武井さん以外は眼球運動な課題。
まあご本人もおっしゃってたけど。
あの流れのなかで、ぽつんと脊髄っていうのは、なかなか異質なカンジでしたね。
muscle synergyのハナシも、あたしはなかなか楽しめましたけどね。
自分の研究と関係するのは、やっぱり最初と最後。
山本先生と橘先生の発表ですかね。
ようするに彦坂研ってことですが。
LS5 神経細胞の活動を見るどうやらそういうことではないようで。
皮質下構造や小脳を対象としてはいるものの、あくまで認知屋な内容です。
というか、3人目の武井さん以外は眼球運動な課題。
まあご本人もおっしゃってたけど。
あの流れのなかで、ぽつんと脊髄っていうのは、なかなか異質なカンジでしたね。
muscle synergyのハナシも、あたしはなかなか楽しめましたけどね。
自分の研究と関係するのは、やっぱり最初と最後。
山本先生と橘先生の発表ですかね。
ようするに彦坂研ってことですが。
大木 研一
3日目はニコンのランチョンセミナーにしまして。
ランチョンが大木先生って、豪華にもほどがあるわけですが。
内容としては、「顕微鏡で何がみえるか」みたいなハナシのみで。
ご自身の研究に関する細かい説明とか、そういうのはほぼナシ。
ズコー。
EL7 教育講演7ランチョンが大木先生って、豪華にもほどがあるわけですが。
内容としては、「顕微鏡で何がみえるか」みたいなハナシのみで。
ご自身の研究に関する細かい説明とか、そういうのはほぼナシ。
顕微鏡のコマーシャルの実用パート
みたいな立ち位置でのトークでした。ズコー。
宮川 剛
マウスの行動バッテリーの第一人者である、宮川先生の教育講演。
席全部埋まって、さらに立ち見多数という盛況ぶりでした。
ひとつひとつの行動課題も、もちろん面白いんですが。
それを数々の遺伝子改変マウスにやってきた、メタデータ的な研究の質量が圧巻で。
そのみちのエキスパートならではの説得力のある内容だった。
その一方で、行動データ解析において、なにに気をつけなければならないか。
種々の「ありがちな」混交要因や、それに対する対策など、実用的な内容も満載で。
まさに教育的な内容だとおもいました。
とくに、さまざまなマウスの行動課題に関して、どんな課題だろうが、
席全部埋まって、さらに立ち見多数という盛況ぶりでした。
ひとつひとつの行動課題も、もちろん面白いんですが。
それを数々の遺伝子改変マウスにやってきた、メタデータ的な研究の質量が圧巻で。
そのみちのエキスパートならではの説得力のある内容だった。
その一方で、行動データ解析において、なにに気をつけなければならないか。
種々の「ありがちな」混交要因や、それに対する対策など、実用的な内容も満載で。
まさに教育的な内容だとおもいました。
とくに、さまざまなマウスの行動課題に関して、どんな課題だろうが、
「その課題で測りたい能力よりも、混交要因のほうが多いぐらいだ」
という説明は、印象的だった。4日目(2012.09.21)
O4-G-62 動機づけ・情動 3、報酬・意思決定 1岡本 仁
あまりに聞くものがないので、またオーラルに入ってみまして。
多少は関係ありそうな、後ろ2人のかたのトコだけ。
しかし、1人目(セッションの3人目)の重宗さんというひとは、どうにも意味わからず。
なんぞグラフは出てくるんだけど。
どういうデータから主張をしてるのかが、わかりませんでした。
脳部位のハナシ以前に、行動データの解釈が意味不明。
2人目の山中航さんは、視床CM核という点以外は、一般的な報酬性意思決定。
なんというか
あたしが「基底核な意思決定業界のハナシ」をフォローしてないせいなんですが。
他の核との関係で、その活動がどういう意味をもつのか、ちとわからなかった。
LS8 最新の顕微鏡イメージングと3次元画像解析多少は関係ありそうな、後ろ2人のかたのトコだけ。
しかし、1人目(セッションの3人目)の重宗さんというひとは、どうにも意味わからず。
なんぞグラフは出てくるんだけど。
どういうデータから主張をしてるのかが、わかりませんでした。
脳部位のハナシ以前に、行動データの解釈が意味不明。
2人目の山中航さんは、視床CM核という点以外は、一般的な報酬性意思決定。
なんというか
「そういう活動でてくるんだー」
みたいな印象で。あたしが「基底核な意思決定業界のハナシ」をフォローしてないせいなんですが。
他の核との関係で、その活動がどういう意味をもつのか、ちとわからなかった。
市川 謙
最終日のランチョンはZeissにしましたが。
講演者は、両方Zeissの社員のかたで。
顕微鏡のコマーシャルと、ソフトの使い方のみ。
すいません、そこまでの内容になっちゃうと、アタシは部外者です。
タダめし食っただけやんけorz
まあ、三次元構造再構築ソフトは、かなり興味深く聞いてたんですがね。
ちょうどいま、ボリュームレンダリングとか勉強してるので。
講演者は、両方Zeissの社員のかたで。
顕微鏡のコマーシャルと、ソフトの使い方のみ。
すいません、そこまでの内容になっちゃうと、アタシは部外者です。
タダめし食っただけやんけorz
まあ、三次元構造再構築ソフトは、かなり興味深く聞いてたんですがね。
ちょうどいま、ボリュームレンダリングとか勉強してるので。
ポスター
1日目(2012.09.18)
P1-g05 サル視床下部外側野における不確かさのコーディング*則武 厚, 中村 加枝
P1-i01 経路探索課題中の前頭前野興奮性-抑制性バランスの時間発展
坂本 一寛, 虫明 元
件のグリッド型迷路をつかった、経路探索課題中のPFCニューロン活動記録。
迷路自体は画面に表示され、サルは手元のレバーかボタンかなんかで自機位置を操作。
神経活動の説明要因は、「最終的なゴール位置」と「初手の方向」の2要因。
波形からニューロンを錐体細胞と介在細胞にわけ、それぞれで解析。
反応に先立ち、興奮・抑制両タイプとも、発火頻度自体はあがっていく。
で、それにつづいて、条件間での発火頻度の差が生じはじめる。
ゴールの表現は、集団全体の動的活動状態でなされてるってことらしい。
P1-i05 エンコード・実行時で異なるニューロン群が反応タクティクスとアクションをコードする*迷路自体は画面に表示され、サルは手元のレバーかボタンかなんかで自機位置を操作。
神経活動の説明要因は、「最終的なゴール位置」と「初手の方向」の2要因。
波形からニューロンを錐体細胞と介在細胞にわけ、それぞれで解析。
反応に先立ち、興奮・抑制両タイプとも、発火頻度自体はあがっていく。
で、それにつづいて、条件間での発火頻度の差が生じはじめる。
ゴールの表現は、集団全体の動的活動状態でなされてるってことらしい。
松坂 義哉, 斎藤 雄太郎, 虫明 元
P1-i11 複数の場所を記憶するための神経機構
松嶋 藻乃, 田中 真樹
ポスター時間帯になっても松嶋さんがいらっしゃらず。
かわりに田中先生にご説明いただいた。
超ラッキー。
基本的には、2ヶ所の空間位置を覚えさせる記憶誘導性眼球運動課題。
手がかり刺激が2ヶ所にでて、両方の位置を覚えておく必要がある。
その刺激が同側に2個か対側に1個ずつかで、続く遅延期の活動が違う、というハナシ。
わたし自身の実験ともメチャメチャ関連してるので、今後にも超注目。
かわりに田中先生にご説明いただいた。
超ラッキー。
基本的には、2ヶ所の空間位置を覚えさせる記憶誘導性眼球運動課題。
手がかり刺激が2ヶ所にでて、両方の位置を覚えておく必要がある。
その刺激が同側に2個か対側に1個ずつかで、続く遅延期の活動が違う、というハナシ。
わたし自身の実験ともメチャメチャ関連してるので、今後にも超注目。
2日目(2012.09.19)
P2-a01 ヒト盲視被験者における視覚誘導性および記憶誘導性サッカード運動吉田 正俊, 雨宮 史織, 伊佐 正
例によって昼前ごろになって現われた吉田先生に説明していただいた。
盲視の協力者さんの眼球運動に関する報告。
生まれつきのV1性片側皮質盲で、物体弁別とかは不可なんだけど。
VGSもMGSもできる。
しかし刺激が存在する場合、サッケードがアンダーシュートする。
アンチサッケードにしても、VGSである限り同じ傾向。
でもMGSだとそれがなくなるという。
なんでだろ…不思議。
P2-a04 線条体ドーパミンの自発性サッカードのタイミングへの関与盲視の協力者さんの眼球運動に関する報告。
生まれつきのV1性片側皮質盲で、物体弁別とかは不可なんだけど。
VGSもMGSもできる。
しかし刺激が存在する場合、サッケードがアンダーシュートする。
アンチサッケードにしても、VGSである限り同じ傾向。
でもMGSだとそれがなくなるという。
なんでだろ…不思議。
國松 淳, 田中 真樹
自分でタイミングをはからなきゃならないMGSをサルに行なわせ。
課題パフォーマンスに対する線条体へのドーパミン薬の効果を調べた実験。
注入はA20~25あたりということで、headとbodyのあいだあたり?
結果としては、D2拮抗薬だけタイミングを前倒しさせる、と。
Motor Control研究会のときにお聞きしたのと、ほぼ同じ結果のままかな?
中村先生の以前の研究とどう対応するのかが難しいし。
エラー率増加による強化率の違いの影響が気になるところだけど。
そのへんの改善は今後、とのこと。
P2-a34 顔の倒立呈示でサル下側頭皮質ニューロンがコードする顔の個体や表情についての情報が減少する*課題パフォーマンスに対する線条体へのドーパミン薬の効果を調べた実験。
注入はA20~25あたりということで、headとbodyのあいだあたり?
結果としては、D2拮抗薬だけタイミングを前倒しさせる、と。
Motor Control研究会のときにお聞きしたのと、ほぼ同じ結果のままかな?
中村先生の以前の研究とどう対応するのかが難しいし。
エラー率増加による強化率の違いの影響が気になるところだけど。
そのへんの改善は今後、とのこと。
菅生-宮本 康子, 河野 憲二
P2-e31 前頭皮質第5層における興奮・抑制性細胞間の結合特性*
森島 美絵子, 川口 泰雄
ラットの前頭皮質5層の錐体細胞を、投射先によって2種に分け。
他の介在細胞との入出力関係を比較。
細かい比率とかはデータが増えればまた変わると思うので。
今後の論文での報告を要チェック。
P2-h17 メチルフェニデート経口投与による前頭連合野と線条体におけるドーパミン放出の増加他の介在細胞との入出力関係を比較。
細かい比率とかはデータが増えればまた変わると思うので。
今後の論文での報告を要チェック。
小島 崇
抄録のほうではauthorはひとりになってるけど。
ポスターのほうでは3名ぐらい名前が載ってて、ラストは渡辺正孝先生。
で、ちょうど渡辺先生がいらっしゃったので、説明していただいた。
ラッキー。
ようするに、ヒトでリタリン飲ませても、脳内のMPH濃度を直接は測れないので。
(PETとか手はなくはないが。)
サルのマイクロダイアリシスでやってみた、と。
基底核のほうでは、そこそこの量の投与でも細胞外DA濃度増加があるが。
5~10mg/kgという大量投与しないと、PFCでのDA増加は検出できない。
ただし、空間性WM課題やGo/No-Go課題の成績改善は少量投薬でもあるので。
PFCで明らかなDA増加が起きなくても、どっかで効いてるのは確か、と。
P2-h20 若年時前頭前野ドーパミン・セロトニン放出とオープンフィールド行動の関係に対する幼弱時手綱核破壊の影響ポスターのほうでは3名ぐらい名前が載ってて、ラストは渡辺正孝先生。
で、ちょうど渡辺先生がいらっしゃったので、説明していただいた。
ラッキー。
ようするに、ヒトでリタリン飲ませても、脳内のMPH濃度を直接は測れないので。
(PETとか手はなくはないが。)
サルのマイクロダイアリシスでやってみた、と。
基底核のほうでは、そこそこの量の投与でも細胞外DA濃度増加があるが。
5~10mg/kgという大量投与しないと、PFCでのDA増加は検出できない。
ただし、空間性WM課題やGo/No-Go課題の成績改善は少量投薬でもあるので。
PFCで明らかなDA増加が起きなくても、どっかで効いてるのは確か、と。
瀧田 正寿, 後藤 幸織
手綱核破壊でDA・5HTシステムを邪魔してやって。
そのときのラットの行動を調べた、と。
1日目のDAや5HT濃度が、1~2日目の運動指標と相関してる、とのこと。
PFC屋は、やっぱDA系の仕事は気になるという雑談をしてたんだけど。
最近はダイアリシスも簡単になったとかおっしゃってたが、ホントだろうか…?
P2-i14 サル背側縫線核における行動選択および価値の表象*そのときのラットの行動を調べた、と。
1日目のDAや5HT濃度が、1~2日目の運動指標と相関してる、とのこと。
PFC屋は、やっぱDA系の仕事は気になるという雑談をしてたんだけど。
最近はダイアリシスも簡単になったとかおっしゃってたが、ホントだろうか…?
林 和子, 則武 厚, 中村 加枝
中村加枝先生が説明なさってたんだけど。
取り巻きがメチャメチャ多くて、ぜんぜん聞こえなかった。
超人気。
内容としては、複数の課題をつかって、行動と報酬価値の異なる状況をつくってやる。
報酬の軸は、ジュースかエアパフか。
行動の軸は、オペラントか古典的条件付けか。
んで、縫線核のニューロン活動を調べた、と。
なんとなくまだ「いろんなニューロンとれてきた」という段階な印象を受けたけど。
十分なデータベースが集まれば、もっといろいろみえてきて面白そう。
しかしコレ、課題条件たくさんあるから。
記録はえらいたいへんそうだな。
P2-j02 背内側線条体から投射する線条体黒質路は弁別課題遂行時の反応時間を調整する*取り巻きがメチャメチャ多くて、ぜんぜん聞こえなかった。
超人気。
内容としては、複数の課題をつかって、行動と報酬価値の異なる状況をつくってやる。
報酬の軸は、ジュースかエアパフか。
行動の軸は、オペラントか古典的条件付けか。
んで、縫線核のニューロン活動を調べた、と。
なんとなくまだ「いろんなニューロンとれてきた」という段階な印象を受けたけど。
十分なデータベースが集まれば、もっといろいろみえてきて面白そう。
しかしコレ、課題条件たくさんあるから。
記録はえらいたいへんそうだな。
深堀 良二, 小林 和人
P2-j14 ラットは系列順序情報を想起したプランニング行動を示した*
石野 誠也, 櫻井 芳雄
ラットに複数の系列反応を訓練。
次のノーズポークホールのLEDが点くから、系列覚えなくても反応はできるんだけど。
訓練により反応時間が短くなるので、実際にはラットは系列を覚えてる、と。
で、その横にたしか神経活動が載ってたとおもうんだけど。
今回、櫻井研のポスターが3枚くらい出てたもんだから。
果たしてそれがこの発表だったかどうか、記憶が定かでない。
なんか、特定の系列中にのみ発火する細胞とか。
系列の進行に伴い、なんとなく活動があがってく細胞とかがあって。
後者のタイプを
はて、このポスターだったかどうか…。
次のノーズポークホールのLEDが点くから、系列覚えなくても反応はできるんだけど。
訓練により反応時間が短くなるので、実際にはラットは系列を覚えてる、と。
で、その横にたしか神経活動が載ってたとおもうんだけど。
今回、櫻井研のポスターが3枚くらい出てたもんだから。
果たしてそれがこの発表だったかどうか、記憶が定かでない。
なんか、特定の系列中にのみ発火する細胞とか。
系列の進行に伴い、なんとなく活動があがってく細胞とかがあって。
後者のタイプを
「系列内での現在の位置(回数)を表現してる」
って書いてるのが、スゲーウソくさいと思った気がするんだが。はて、このポスターだったかどうか…。
3日目(2012.09.20)
P3-a09 背外側線条体から投射する線条体淡蒼球路は聴覚弁別学習の正確な遂行を調節する*西澤 佳代, 小林 和人
P3-a32 二光子カルシウムイメージングのためのマウス仮想ナビゲーションシステム*
佐藤 正晃, 林 康紀
P3-a33 視覚刺激と聴覚刺激の時間弁別におけるサル前頭前野の神経細胞活動*
千葉 惇, 生塩 研一, 稲瀬 正彦
P3-a38 ラット海馬CA1におけるエラー試行時の局所脳波解析
西田 洋司, 高橋 宗良, Lauwereyns Johan
2ヶ所のノーズポークホールでの交替反応課題をラットに訓練し。
その間のCA1のLFPやスパイクを解析した研究。
課題関連の周波数解析に加え、シータ帯位相とスパイクとのロックの解析が面白い。
ポークホール選択性細胞は、preferred反応時に位相とのロックがうしろにズレていくが。
non-preferredなホールへの反応時には、それがない。
しかもその位相とのズレ方は、早期誤反応試行では急峻になるという。
高ガンマ帯とシータ帯にわけることによる、信号ソースの考察も興味深かった。
CA3性かentorhinal性かっていう。
まあ、entorhinalからの情報って、内容がなんでもアリすぎてよくわからないが。
そのへんは今後、ってとこでしょうか。
P3-b01 カニクイザル頭蓋からの脳区分を推定する*その間のCA1のLFPやスパイクを解析した研究。
課題関連の周波数解析に加え、シータ帯位相とスパイクとのロックの解析が面白い。
ポークホール選択性細胞は、preferred反応時に位相とのロックがうしろにズレていくが。
non-preferredなホールへの反応時には、それがない。
しかもその位相とのズレ方は、早期誤反応試行では急峻になるという。
高ガンマ帯とシータ帯にわけることによる、信号ソースの考察も興味深かった。
CA3性かentorhinal性かっていう。
まあ、entorhinalからの情報って、内容がなんでもアリすぎてよくわからないが。
そのへんは今後、ってとこでしょうか。
小林 靖, 松村 讓兒
「なんでそんなことを」っておもったが。
化石人類のキャストから脳の発達を調べるとか、そういう目的性らしく。
その方法論を、原生霊長類で模索してるってことなのね。
しかし、化石なんていろんな侵食作用にさらされてるわけだから。
骨からの脳容積推定手法がどれだけ化石にも適用可能かは、ようわからん。
P3-c04 マーモセット大脳皮質3a野の神経生理学と解剖学的研究*化石人類のキャストから脳の発達を調べるとか、そういう目的性らしく。
その方法論を、原生霊長類で模索してるってことなのね。
しかし、化石なんていろんな侵食作用にさらされてるわけだから。
骨からの脳容積推定手法がどれだけ化石にも適用可能かは、ようわからん。
纐纈 大輔, 伊佐 正, 南部 篤
P3-c05 ヒゲ感覚システムにおける大脳皮質深部の遠距離投射ニューロンが形成する回路の構造と機能*
平井 大地, 古田 貴寛, 金子 武嗣
金子先生のところのこういう仕事、すごいなとはおもうんだけど。
どれだけこの結果が一般化できるかってトコが、いつも疑問なんだよね。
もちろん丹念に投射先を調べるってのは、それはそれで重要だとおもうが。
10個以下みたいなサンプルサイズだと…ね。
P3-c11 マウス大脳皮質における後肢体性感覚地図*どれだけこの結果が一般化できるかってトコが、いつも疑問なんだよね。
もちろん丹念に投射先を調べるってのは、それはそれで重要だとおもうが。
10個以下みたいなサンプルサイズだと…ね。
井上 舞, 鈴木 崇之, 村山 正宜
麻酔したマウスの後肢の各部位を感覚刺激し。
そのときのS1の誘発応答をVSDでマッピングしたという報告。
なんでそもそも後肢でやろうとおもったのかが、ちょっと疑問だが…。
誘発応答もかなりひろく出てて、体性局在もわかりにくいし。
結局、ゆるーい手のひらの絵を、皮質上にマンガで描くぐらいのマップになっていた。
前肢とか、あるいはそれこそヒゲとかでやれば、もっと綺麗に出るのは明らかなんだが。
それはもうやられちゃってるから、残ってる後肢で、っていうことなのかな?
P3-d14 ラットのタイミング行動課題中における時間知覚と小脳プルキンエ細胞のスパイク発火そのときのS1の誘発応答をVSDでマッピングしたという報告。
なんでそもそも後肢でやろうとおもったのかが、ちょっと疑問だが…。
誘発応答もかなりひろく出てて、体性局在もわかりにくいし。
結局、ゆるーい手のひらの絵を、皮質上にマンガで描くぐらいのマップになっていた。
前肢とか、あるいはそれこそヒゲとかでやれば、もっと綺麗に出るのは明らかなんだが。
それはもうやられちゃってるから、残ってる後肢で、っていうことなのかな?
山口 健治, 高橋 晋, 櫻井 芳雄
DRLで複数回タッチパネルに前肢でタッチする課題をラットに訓練し。
その間の小脳プルキンエ細胞の活動を取ったという研究。
時間表現なので小脳というのはまあわかるんだけど。
DRL3秒とかの反応は、果たしてどのレベルの時間情報処理になるのか。
タイミング行動とか運動協調みたいなレベルなら、小脳でいいんだろうけど。
経時のようなintermediate timeに興味があるなら、皮質のほうがよくね?
課題関連応答も(いまのとこ)面白いのいなかったっていってたし。
どうでもいいが、発表者のひとと雑談してて。
生塩先生の実験のハナシしてたら、うしろに御本人がいらっしゃって超びびった(笑)
あちらは時間弁別課題なので、DRLとはみてるものがちょっと違うけどね。
P3-e32 新規光刺激装置による自由行動下マウスの運動制御その間の小脳プルキンエ細胞の活動を取ったという研究。
時間表現なので小脳というのはまあわかるんだけど。
DRL3秒とかの反応は、果たしてどのレベルの時間情報処理になるのか。
タイミング行動とか運動協調みたいなレベルなら、小脳でいいんだろうけど。
経時のようなintermediate timeに興味があるなら、皮質のほうがよくね?
課題関連応答も(いまのとこ)面白いのいなかったっていってたし。
どうでもいいが、発表者のひとと雑談してて。
生塩先生の実験のハナシしてたら、うしろに御本人がいらっしゃって超びびった(笑)
あちらは時間弁別課題なので、DRLとはみてるものがちょっと違うけどね。
橋本 光広, 平瀬 肇, 宮田 卓樹
LEDの刺激装置をChR2発現マウスの運動野(?)に埋め込んでおき。
制御はあたまに固定した通信装置を介して、ワイヤレス。
これで片側の皮質を刺激すると、それと逆側にマウスの進行方向を曲げられた、という。
バレル刺激してるんじゃないかという気もするんだけど、M1でいいのかな?
P3-e34 光遺伝学的刺激により引き込まれたラット大脳皮質の除波振動のアップ状態の高周波活動の解析制御はあたまに固定した通信装置を介して、ワイヤレス。
これで片側の皮質を刺激すると、それと逆側にマウスの進行方向を曲げられた、という。
バレル刺激してるんじゃないかという気もするんだけど、M1でいいのかな?
九鬼 敏伸, 八尾 寛, 虫明 元
睡眠や麻酔下での皮質応答には、up stateとdown stateがあらわれる。
で、stateごとに優位な周波数帯が違い、しかもそれがstate内で遷移していく。
そこで、ChR2発現マウスの皮質に光刺激で無理やり特定の周波数入力を行ない。
それによって人工的にstateを切り替えできるんだそうな。
あたしにはそれがどういう役に立つかイマイチわからないが、単純に面白い。
P3-e35 光遺伝学と逆行性遺伝子導入を利用した大脳皮質-線条体経路選択的な興奮誘導で、stateごとに優位な周波数帯が違い、しかもそれがstate内で遷移していく。
そこで、ChR2発現マウスの皮質に光刺激で無理やり特定の周波数入力を行ない。
それによって人工的にstateを切り替えできるんだそうな。
あたしにはそれがどういう役に立つかイマイチわからないが、単純に面白い。
佐野 裕美, 小林 和人, 南部 篤
投射元と投射先からそれぞれ順向性・逆向性ウイルスベクタを入れてやることで。
特定の投射ニューロンにだけChR2を発現させて刺激できたという実験。
こういうの、ホントいいですねー。
霊長類でもこういう方法論がどんどん発達してほしい。
回路機能を直接的に調べる意味で、桁外れに強力な手法だとおもうので。
特定の投射ニューロンにだけChR2を発現させて刺激できたという実験。
こういうの、ホントいいですねー。
霊長類でもこういう方法論がどんどん発達してほしい。
回路機能を直接的に調べる意味で、桁外れに強力な手法だとおもうので。
4日目(2012.09.21)
P4-d16 高分解能MRIによるマーモセット大脳の脳溝形成の解析*澤田 和彦, 岡野 栄之
P4-f13 ラット感覚運動学習に対する強制応答動作の影響*
金子 秀和, 鈴木 慎也
左右のレバーと手がかり刺激をつかったレバー離し課題をラットに訓練。
その際、無理やりレバーをあげてリリース運動を誘発させる強制動作条件をつくってみたが。
学習にはマイナスにはたらいてしまったという結果。
発想としては、たぶん無誤学習みたいなハナシで。
正反応の経験を無理やりさせてやれば、学習を促進するだろう、ってことだとおもう。
なんでダメだったんだろうね。
やっぱ動物が随意的に反応する余地がないと、オペラント学習ができないってコトだろうか。
P4-f14 感覚運動学習における早い学習過程と遅い学習過程の神経基盤その際、無理やりレバーをあげてリリース運動を誘発させる強制動作条件をつくってみたが。
学習にはマイナスにはたらいてしまったという結果。
発想としては、たぶん無誤学習みたいなハナシで。
正反応の経験を無理やりさせてやれば、学習を促進するだろう、ってことだとおもう。
なんでダメだったんだろうね。
やっぱ動物が随意的に反応する余地がないと、オペラント学習ができないってコトだろうか。
今水 寛, Nicolas Schweighofer
ヒトでカーソル移動反応にperturbationを与えてやって。
その運動学習過程を、30段階ぐらいの異なる時定数成分をもつstate modelで推定。
ついでにその間の脳活動から、責任部位を同定したという実験。
結果、早い成分は高次運動野や頭頂連合野の活動に帰され。
遅い成分になるにつれ、小脳性の寄与が強くなったということらしい。
あたしは門外漢なんで、詳しい計算過程はぜんぜんわからないが。
課題のつくりから解析までが理路整然としていて、すごくわかりやすかった。
P4-f16 視覚運動変換の適応において運動終了と課題終了はいずれも誤差フィードバックにとって重要なタイミングである*その運動学習過程を、30段階ぐらいの異なる時定数成分をもつstate modelで推定。
ついでにその間の脳活動から、責任部位を同定したという実験。
結果、早い成分は高次運動野や頭頂連合野の活動に帰され。
遅い成分になるにつれ、小脳性の寄与が強くなったということらしい。
あたしは門外漢なんで、詳しい計算過程はぜんぜんわからないが。
課題のつくりから解析までが理路整然としていて、すごくわかりやすかった。
石川 拓海, 阪口 豊
P4-f17 運動準備期間の脳活動を用いた誤差の予測*
大畑 龍, 今水 寛
ヒトに指運動課題をさせ、その間の脳活動をMEGで記録。
指運動の誤差のでかい試行と小さい試行をとってきて、MEG信号から両条件を機械学習。
結果、運動前野の信号からの予測性がとくに強かった。
P4-f25 VBMEGによる運動準備電位を定位する*指運動の誤差のでかい試行と小さい試行をとってきて、MEG信号から両条件を機械学習。
結果、運動前野の信号からの予測性がとくに強かった。
蔡 暢, 今水 寛
P4-f26 ニホンザル運動関連領野のフラビンタンパク蛍光イメージング
畑中 伸彦, 南部 篤
内因性のフラビン蛍光をサルの運動皮質でイメージングしてやろうという研究。
ただし、やはりduraをはさむとぜんぜん信号返ってこなかったんで。
duraを除去したうえからシリコン性の人工硬膜をあて。
そのあと浮腫などの急性反応をひととおり治療してからイメージングしたという。
ようやるな、そんなたいへんなこと。
結果的には、ボタン押し課題でかなりちゃんとした信号取れて。
内転-外転(内旋-外旋か?)の軸での皮質応答の差や。
同側運動時の高次運動野に限局した応答も取れた、と。
ただし、人工硬膜の内側に組織がはるせいで、やはりレコーディングの水準は低下するそうで。
4ヶ月くらい経つと、弱めの信号は検出がむずかしくなってくるとか。
P4-f27 サルが円状の閉曲線を描く際の経由点の推定*ただし、やはりduraをはさむとぜんぜん信号返ってこなかったんで。
duraを除去したうえからシリコン性の人工硬膜をあて。
そのあと浮腫などの急性反応をひととおり治療してからイメージングしたという。
ようやるな、そんなたいへんなこと。
結果的には、ボタン押し課題でかなりちゃんとした信号取れて。
内転-外転(内旋-外旋か?)の軸での皮質応答の差や。
同側運動時の高次運動野に限局した応答も取れた、と。
ただし、人工硬膜の内側に組織がはるせいで、やはりレコーディングの水準は低下するそうで。
4ヶ月くらい経つと、弱めの信号は検出がむずかしくなってくるとか。
鍋 知宏, 宮下 英三
P4-f31 ドーパミンD1受容体欠損およびドーパミンD2受容体欠損マウスの運動量の詳細解析*
佐藤 朝子, 笹岡 俊邦
P4-f33 運動の実行・非実行に関わるラット一次・二次運動野のマルチニューロン活動*
齊木 愛希子, 礒村 宜和
このあいだの玉川のプログラムでお世話になった、磯村先生のところの齊木さんの発表。
みにこないよう釘をさされたので、説明に立ってないときを見計らってこっそり読みにいった。
頭固定のラットにGo/No-Go課題を訓練し。
おなじ「引き」動作時の神経活動を比較。
さらにそれを、M1とM2間で比べてみたという研究。
とりあえずざっとみたカンジだと、あんまり皮質間で差はないのかな?
cross-correlationのほうも、一貫した面白い傾向ってのは出てないみたい。
multiで取ってると、やっぱ強みは細胞間相関の解析ができることだけど。
2細胞間だけでもいろんなパターンがあって、ストーリーまとめるのが難しいのよね。
各細胞のPSTH特性と細胞間の相関関係をどうまとめるか。
そのへんはこれからってカンジなんでしょうか。
それにしても、No-Go試行での一番重要なイベントは、やっぱNo-Goの瞬間だと思うんだけど。
そこの解析はもっと詳しくしなくていいのかな…?
運動条件をそろえて試行条件間比較をするってのは、まあわかるんだけどさ。
あと、No-Go時の運動をもうちょっと細かく解析するとか。
いろいろこれからの発展はありそうな。
P4-f34 皮質発火時系列の構造が示唆する最適な発火率カーネルによる経過時間コーディングみにこないよう釘をさされたので、説明に立ってないときを見計らってこっそり読みにいった。
頭固定のラットにGo/No-Go課題を訓練し。
おなじ「引き」動作時の神経活動を比較。
さらにそれを、M1とM2間で比べてみたという研究。
とりあえずざっとみたカンジだと、あんまり皮質間で差はないのかな?
cross-correlationのほうも、一貫した面白い傾向ってのは出てないみたい。
multiで取ってると、やっぱ強みは細胞間相関の解析ができることだけど。
2細胞間だけでもいろんなパターンがあって、ストーリーまとめるのが難しいのよね。
各細胞のPSTH特性と細胞間の相関関係をどうまとめるか。
そのへんはこれからってカンジなんでしょうか。
それにしても、No-Go試行での一番重要なイベントは、やっぱNo-Goの瞬間だと思うんだけど。
そこの解析はもっと詳しくしなくていいのかな…?
運動条件をそろえて試行条件間比較をするってのは、まあわかるんだけどさ。
あと、No-Go時の運動をもうちょっと細かく解析するとか。
いろいろこれからの発展はありそうな。
坪 泰宏, 深井 朋樹
こちらは、以前の磯村先生のデータをもとにした理論寄りの研究。
観察されたニューロンのISIヒストグラムの大半が、power lawだということから。
それを経過時間の表現とからめて議論したというものらしい。
あたしは数理詳しくないから、あまり細かいことはわからんくて…。
ISIヒストグラムとdensity functionは、どういう関係になるのかな?
発火率カーネルは後者のハナシだよね?
P4-f35 一次運動野と運動前野は視覚エラーと非視覚エラーをコードしている観察されたニューロンのISIヒストグラムの大半が、power lawだということから。
それを経過時間の表現とからめて議論したというものらしい。
あたしは数理詳しくないから、あまり細かいことはわからんくて…。
ISIヒストグラムとdensity functionは、どういう関係になるのかな?
発火率カーネルは後者のハナシだよね?
井上 雅仁, 内村 元昭, 北澤 茂
プリズムによるperturbation入りの到達課題をサルに行わせ。
そのときの運動野ニューロンによるエラー表象を調べた実験。
面白いんだが、文章では視覚・非視覚エラーの違いを説明しづらいorz
P4-f36 手指の停止動作に対する経頭蓋磁気刺激による修飾そのときの運動野ニューロンによるエラー表象を調べた実験。
面白いんだが、文章では視覚・非視覚エラーの違いを説明しづらいorz
樋脇 治, 石丸 尚之, 福田 浩士
4秒に一回の2Nでのボタン押しを被験者にやらせるが。
50%の試行でストップシグナルを入れる。
そのストップの前後のいろんなタイミングで、運動野にTMSを打ったというもの。
結果、ストップ直前のTMSは運動抑制を助けるはたらきをし(発揮力の減少)。
逆にストップシグナル直後のTMSは運動抑制を阻害した(発揮力増加)。
TMSは丸形コイルで広範にうってるから、責任部位はハッキリしないが。
大脳縦裂でSMAに強く作用して、こういう結果になったんじゃないか、とのこと。
個人的に、ストップ試行が50%というのはかなり多いと思うので。
それについて質問したんだが。
ストップ試行の割合は結果に関係しないだろうとのことで。
う~ん…そうですかねぇ?
実際、RTバージョンでストップ試行率変えると、RT分布変わるんで。
それって、ストップの予測性に依存して、準備状態が違うってことでしょ。
だとしたら、TMSの影響がかわってもおかしくないとおもうんだけどなぁ。
たとえば、プラス-マイナス効果の切り替わりのタイムコースがかわるとか。
P4-l04 ラット自由行動下における内側前頭前皮質のマルチユニット活動*50%の試行でストップシグナルを入れる。
そのストップの前後のいろんなタイミングで、運動野にTMSを打ったというもの。
結果、ストップ直前のTMSは運動抑制を助けるはたらきをし(発揮力の減少)。
逆にストップシグナル直後のTMSは運動抑制を阻害した(発揮力増加)。
TMSは丸形コイルで広範にうってるから、責任部位はハッキリしないが。
大脳縦裂でSMAに強く作用して、こういう結果になったんじゃないか、とのこと。
個人的に、ストップ試行が50%というのはかなり多いと思うので。
それについて質問したんだが。
ストップ試行の割合は結果に関係しないだろうとのことで。
う~ん…そうですかねぇ?
実際、RTバージョンでストップ試行率変えると、RT分布変わるんで。
それって、ストップの予測性に依存して、準備状態が違うってことでしょ。
だとしたら、TMSの影響がかわってもおかしくないとおもうんだけどなぁ。
たとえば、プラス-マイナス効果の切り替わりのタイムコースがかわるとか。
南 千尋, 三谷 章
P4-m12 視覚探索におけるサル後頭頂葉の目標刺激選別過程は刺激の明るさおよび目標刺激と妨害刺激の色の違いにより異なる影響を受ける
田中 智洋, 西田 知史, 小川 正
LIPニューロンは、眼球運動のターゲットがRFに入ったとき、発火頻度が漸増するが。
その活動増加の文脈依存性を調べた報告。
6肢のうちひとつだけ色が違うsingletonのVGSをサルに訓練し、LIPニューロンを記録。
このとき、手がかり刺激の色の全体的な明るさで2条件(light/dark)あり。
暗いほうがより難しい。
また各明るさ条件内で、さらに難しさの2条件(easy/difficult)があり。
ターゲットと妨害刺激のコントラストが小さいほうが難しい。
で、「lightのdifficult条件」と「darkのeasy条件」は、同程度の難易度となり。
運動出力たる反応時間では差がないのに。
ターゲット位置がRF内・外の試行間でのdifferentialな応答の潜時が異なる。
この活動がどういうことを意味してるのか、解釈はいろいろだとおもうが。
単純にこういう活動の差がでてくるっては、おもしろいね。
それにしても、なぜかポスターには、小川先生の名前しか載ってなかった気がするが。
これって先生がご自身で準備されたポスターなのかな?
P4-m17 視覚探索中の目標刺激の選別過程における,後頭頂葉ニューロンの活動促進・抑制のダイナミクスその活動増加の文脈依存性を調べた報告。
6肢のうちひとつだけ色が違うsingletonのVGSをサルに訓練し、LIPニューロンを記録。
このとき、手がかり刺激の色の全体的な明るさで2条件(light/dark)あり。
暗いほうがより難しい。
また各明るさ条件内で、さらに難しさの2条件(easy/difficult)があり。
ターゲットと妨害刺激のコントラストが小さいほうが難しい。
で、「lightのdifficult条件」と「darkのeasy条件」は、同程度の難易度となり。
運動出力たる反応時間では差がないのに。
ターゲット位置がRF内・外の試行間でのdifferentialな応答の潜時が異なる。
この活動がどういうことを意味してるのか、解釈はいろいろだとおもうが。
単純にこういう活動の差がでてくるっては、おもしろいね。
それにしても、なぜかポスターには、小川先生の名前しか載ってなかった気がするが。
これって先生がご自身で準備されたポスターなのかな?
西田 知史, 田中 智洋, 小川 正
おなじく小川先生のところのLIPのニューロン活動記録。
singletonのVGSで、ターゲット位置がRFとnon-RFの試行の活動を比べるだけでなく。
ターゲットが存在しないキャッチ試行での活動も記録。
それを基準とすると、non-RF試行での活動は、ベースラインより下がった状態になってる。
つまり、他のポピュレーションから抑制されてる、という主張。
個人的には、そんなこと昔から知られてるじゃんとおもうし。
やるんならmultiで取ってcross-correlationとかやるべきな気がするけど。
あと、ポスター内容とは関係ないんですが。
こんな明らさまなとこで、無防備にポスター説明してもらってたりしたので。
まんまと小川先生にみつかって連行されましたorz
singletonのVGSで、ターゲット位置がRFとnon-RFの試行の活動を比べるだけでなく。
ターゲットが存在しないキャッチ試行での活動も記録。
それを基準とすると、non-RF試行での活動は、ベースラインより下がった状態になってる。
つまり、他のポピュレーションから抑制されてる、という主張。
個人的には、そんなこと昔から知られてるじゃんとおもうし。
やるんならmultiで取ってcross-correlationとかやるべきな気がするけど。
「なぜこういう仕事を、このご時世にユニットで?」
と思わないでもない。あと、ポスター内容とは関係ないんですが。
こんな明らさまなとこで、無防備にポスター説明してもらってたりしたので。
まんまと小川先生にみつかって連行されましたorz
感想
まあとりあえずの感想として。4日間は長ぇ。
そんなにラボを開けちゃうと、正直実験がかなり痛手を食うんで。
(出張後のメンテと実験再稼働にかかる時間も含めて。)
はっきりいって、ご遠慮申し上げたいレベルです。
4日間になったからそのぶん内容が充実したかというと、そうでもなく。
どちらかというと、単純に濃度が薄まったカンジで。
みるべきポスターも聞くべきシンポもない空き時間が、けっこう多かった。
加えて、なんか今年は絶対量としても、シンポジウムが不作な年で。
あ、これはあくまで「あたしにとって」ですが。
自分が関係する分野のシンポが、ホント少なかった。
プレナリーと特別講演に関しては、なんと聞きたいのがひとつもないというありさま。
この業界全体がオワコンってことは、ないとおもうんだけどなぁ…。
とはいえ、ヒマが多いってのはわるいことばかりではなくて。
例年よりじっくりとポスターをまわれた気がします。
シンポの開始時間を気にしたりする必要がなかったからね。
いい加減、学会参加も慣れてきて、スケジューリングがマシになったとか。
説明をお願いするのがうまくなってきたとか。
そういう要因もあるかもしれないですが。
しかし、やっぱ自分のポスターのまわりが、一番聞きたい分野の発表なんで。
自分のヤツ説明してると、必然的にそれらをみにいけないってのは、変わりなく。
今年もかなり気になるポスターを、だいぶ見逃しました。
他の日にどんなにヒマがあろうとも、自分の発表日だけはずっと忙しいからね。
これは構造上の問題なんで、大会が何日間になろうと、カンケーないでしょう。
そこで、ふとおもったんだけど。
それを見越して、ちょっと違う分野で演題登録しとくってのも手ですね。
もちろん全然違う分野に出すわけにはいかないけど。
たとえば「注意・知覚統合」でなく「視覚」で出すとか。
吉田先生とかたいてい「視覚」だから、ボッチになることもないだろうし。
来年は、ちと一考に値するな…。