@kanri_ninjin
@kanri_ninjin


 イクナイ! 581
イクナイ! 581







第33回 日本神経科学大会(2010)の記録
2010年の神経科学学会に参加した際の記録です。独断と偏見に基づいた完全なる個人の感想なので、それがイヤなかたはお読みにならないよう、お願いいたします。
ポスターにおける発表者が複数いる場合には、ファーストとラスト、あるいはラストに準ずるひと2~3人だけ抜粋して記載しています。
それから、アスタリスクつきのポスターは、自分でちょろっと読んだだけのヤツ。
アスタリスクなしは、ちゃんと発表者に説明してもらったヤツです。
基本情報
今大会は、日本神経化学学会と日本神経回路学会との、3会合同大会。そのため、名称も「Neuroscience 2010」ではなく、「Neuro2010」
神経科学学会のチェアーは川人光男先生。
どうでもいいけど、今大会から(?)抄録集を紙媒体で配る(または売る)システムはなくなったらしい。
だとすると、抄録集にあたるものは、あとでPDFかなんかでダウンロードできるようにしてくれるのかな?
オンラインの検索システムだけだと、いつまで保守されるか、不安なんだよね。
スケジュール
最終的な、3日間のあたしの動きはこんなカンジ。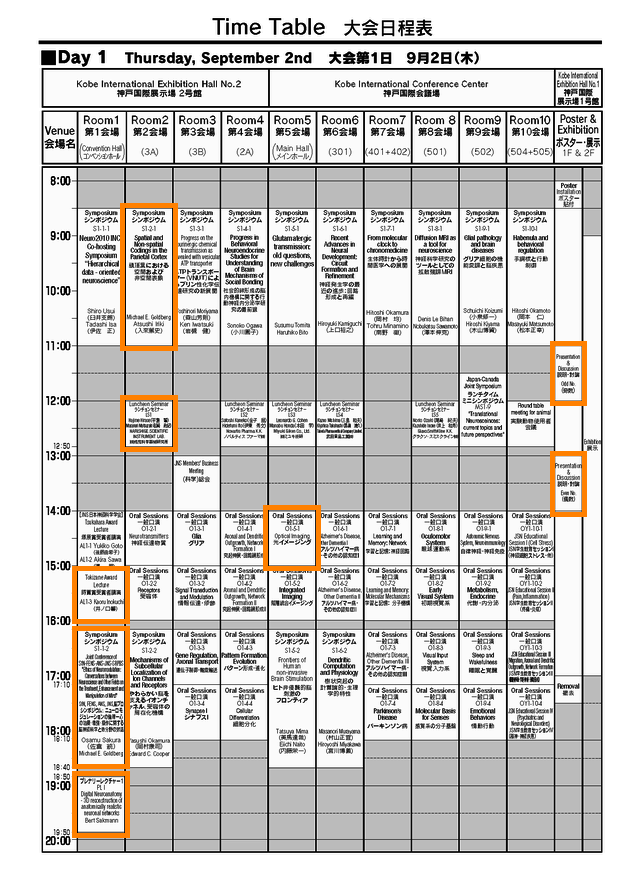


トーク
1日目(2010.09.02)
S1-2-1 頭頂葉における空間および非空間表象Michael E. Goldberg, 入來 篤史
頭頂葉といえば視空間認知。
しかし進化的にみると、頭頂葉は前頭葉に先立って、霊長類で大きく発達した連合皮質。
よって、空間表象以外の役割もあるんじゃね?っていう内容。
最初の入来先生の導入と、Goldbergのマニアックなdouble saccadeのハナシはたいへん面白かったのだが…
そのあとの、件のサル版WCSTのハナシと意図・気付きのハナシは、申し訳ないことにえらい眠かった。
LS1 電気・光生理学によるニューロン・グリアの機能・構造連関研究しかし進化的にみると、頭頂葉は前頭葉に先立って、霊長類で大きく発達した連合皮質。
よって、空間表象以外の役割もあるんじゃね?っていう内容。
最初の入来先生の導入と、Goldbergのマニアックなdouble saccadeのハナシはたいへん面白かったのだが…
そのあとの、件のサル版WCSTのハナシと意図・気付きのハナシは、申し訳ないことにえらい眠かった。
松崎 政紀
初日ランチョンセミナー。
まずはじめに、平瀬先生の、ラボ立ち上げからこれまでの研究について。
つぎに松崎先生の、おもにケージド化合物をつかったsubcellularなイメージングの研究について。
どっちも面白かったが、とくに平瀬先生の、empty roomを与えられてから研究を始動するまでのハナシがよかった。
O1-5-1 光イメージングまずはじめに、平瀬先生の、ラボ立ち上げからこれまでの研究について。
つぎに松崎先生の、おもにケージド化合物をつかったsubcellularなイメージングの研究について。
どっちも面白かったが、とくに平瀬先生の、empty roomを与えられてから研究を始動するまでのハナシがよかった。
川上 浩一, 八田 公平
塚原賞記念講演は知らない方々だったので、時實までのあいだ、オーラルに入ってみた。
で、「オーラルはもういいや」と思い知らせていただいた。
去年も、初日に聞いたオーラルセッションで同じことを思って、以降、オーラルはひとつも聞かなかったんだが…
今年も同じ結末になった。
オーラルセッションってさ、英語でやる意味あるのだろうか?
いやさ、院生に英語プレゼンを経験させる教育の場としては、重要だと思うんだけど。
「原稿を機械音声に読ませました」みたいな音読聞いてても、ぜんぜん内容が聞き取れねぇ。
日本語でやれば、もっと中身のある時間を過ごせると思うのだけど。
あと、緊張してるのだとは思うが、発表する側はもう少しまじめに準備してくるべき。
やるからには、自分が話す内容くらい寝ててもスラスラでるくらいリハーサルしてきてほしい。
AL1-3 長期記憶形成の分子・細胞機構で、「オーラルはもういいや」と思い知らせていただいた。
去年も、初日に聞いたオーラルセッションで同じことを思って、以降、オーラルはひとつも聞かなかったんだが…
今年も同じ結末になった。
オーラルセッションってさ、英語でやる意味あるのだろうか?
いやさ、院生に英語プレゼンを経験させる教育の場としては、重要だと思うんだけど。
「原稿を機械音声に読ませました」みたいな音読聞いてても、ぜんぜん内容が聞き取れねぇ。
日本語でやれば、もっと中身のある時間を過ごせると思うのだけど。
あと、緊張してるのだとは思うが、発表する側はもう少しまじめに準備してくるべき。
やるからには、自分が話す内容くらい寝ててもスラスラでるくらいリハーサルしてきてほしい。
井ノ口 馨
井ノ口先生ってどんなかたか、よく把握せずに聴きにいったが。
シナプスタギングのひとでしたか。
農学部を出て、微生物での分子生物学研究を経たのち、脳の研究に移られたという経歴にびっくりした。
塚原仲晃先生の『脳の可塑性と記憶』を読んで、この道に入ることを突如決意されたとか。(時實賞なのにww)
なんか、そういうひとってけっこう多い気がする。
塚原仲晃は、日本版シュレディンガー先生なのだなあ。
S1-1-2 ニューロモジュレーションの倫理 - 心の治療・増強・操作に関する脳神経科学と他分野の対話シナプスタギングのひとでしたか。
農学部を出て、微生物での分子生物学研究を経たのち、脳の研究に移られたという経歴にびっくりした。
塚原仲晃先生の『脳の可塑性と記憶』を読んで、この道に入ることを突如決意されたとか。(時實賞なのにww)
なんか、そういうひとってけっこう多い気がする。
塚原仲晃は、日本版シュレディンガー先生なのだなあ。
佐倉 統, Michael E. Goldberg
失礼ながら、かなり寝てしまった。
もともと、このあとのSakmannのプレナリーまで時間があるので、入っただけだったし。
しかも発表の多くが、倫理学や哲学の方のトークで、延々と文字だけが書かれているスライドに滅入ってしまって。
PL1 Digital Neuroanatomy - 3D reconstruction of anatomically realistic neuronal networksもともと、このあとのSakmannのプレナリーまで時間があるので、入っただけだったし。
しかも発表の多くが、倫理学や哲学の方のトークで、延々と文字だけが書かれているスライドに滅入ってしまって。
Bert Sakmann
感動した。
研究対象にしてるのは、ネズミのヒゲによる探索行動。
そして、それに必要とされる神経回路のなかでも、視床とバレルというごく一部分だけに注目しているのだが。
コネクションのトボグラフィーを調べ、層ごとの細胞体・樹状突起分布を調べ、細胞形態を調べ、subcellularなマッピングを調べ…
さらにはそれらの実験結果から、ひとつのバレル全体をコンピュータ上に再構築し、細胞種特異的にインプットを与えたときに観察されるcalcium transientをシミュレートする。
ひとつひとつの仕事を、丁寧に、定量的に行なうことによって、それらの総体としてみえてくる神経回路の挙動が美しい。
あと、声に惚れた。
研究対象にしてるのは、ネズミのヒゲによる探索行動。
そして、それに必要とされる神経回路のなかでも、視床とバレルというごく一部分だけに注目しているのだが。
コネクションのトボグラフィーを調べ、層ごとの細胞体・樹状突起分布を調べ、細胞形態を調べ、subcellularなマッピングを調べ…
さらにはそれらの実験結果から、ひとつのバレル全体をコンピュータ上に再構築し、細胞種特異的にインプットを与えたときに観察されるcalcium transientをシミュレートする。
ひとつひとつの仕事を、丁寧に、定量的に行なうことによって、それらの総体としてみえてくる神経回路の挙動が美しい。
あと、声に惚れた。
2日目(2010.09.03)
S2-6-1 小規模回路から探る脳の情報処理基盤能瀬 聡直, 細谷 俊彦
自分のポスターを貼ってから、まだひとが少ないようだったので、シンポジウムに参加。
今大会のシンポのなかでも、密かに期待してたもののひとつ。
カルシウムイメージングやオプトジェネティクスを使って、神経回路の情報処理過程にどのようにアプローチするかというもの。
個人的には、最初の風間先生と最後のChichilnisky先生のハナシがとくに面白かった。
「技術的進歩のためにより複雑な対象も扱えるようになった(それでもなお)単純系アプローチ」としての小規模回路研究って、ほんとに魅力的だと思う。
S2-1-2 負情動の生物学的基盤:生得的回路と獲得的回路今大会のシンポのなかでも、密かに期待してたもののひとつ。
カルシウムイメージングやオプトジェネティクスを使って、神経回路の情報処理過程にどのようにアプローチするかというもの。
個人的には、最初の風間先生と最後のChichilnisky先生のハナシがとくに面白かった。
「技術的進歩のためにより複雑な対象も扱えるようになった(それでもなお)単純系アプローチ」としての小規模回路研究って、ほんとに魅力的だと思う。
加藤 総夫, 喜田 聡
ひとりめのLeDouxだけ聴きに。
LeDouxのプレナリーは聴かずに帰るつもりだったので、声だけでも聞いとこうと思って、という不純な動機。
自分のポスターでヘロヘロだったもので。
スミマセン。
ちなみにこの日はランチョンにもいけずじまいだったため、参加できたシンポが少なくなってしまいました。
LeDouxのプレナリーは聴かずに帰るつもりだったので、声だけでも聞いとこうと思って、という不純な動機。
自分のポスターでヘロヘロだったもので。
スミマセン。
ちなみにこの日はランチョンにもいけずじまいだったため、参加できたシンポが少なくなってしまいました。
3日目(2010.09.04)
S3-10-1 神経信号データの解析法篠本 滋, Sonja Gruen
密かに期待していたシンポその2。
思っていたとおり面白かった。
基本は、マルチニューロンのシグナルをどう解析するかっていうハナシ。
ただし篠本先生だけは、そもそものスパイク系列を可視化・解釈するときの問題など、ベーシックな内容。
たいへん分かりやすい。
あとMoshe Abelesは、神経生理屋なんだけど。
記録技術の進歩のなかで、実験者側の解析方法の洗練の必要を感じ、この歳でHidden Markov Modelsとか勉強したって言ってて感動した。
LS13 研究者と企業のコラボレーションが創出する応用脳科学の世界思っていたとおり面白かった。
基本は、マルチニューロンのシグナルをどう解析するかっていうハナシ。
ただし篠本先生だけは、そもそものスパイク系列を可視化・解釈するときの問題など、ベーシックな内容。
たいへん分かりやすい。
あとMoshe Abelesは、神経生理屋なんだけど。
記録技術の進歩のなかで、実験者側の解析方法の洗練の必要を感じ、この歳でHidden Markov Modelsとか勉強したって言ってて感動した。
川人 光男
ややあやしい(?)ランチョンセミナー。
これからの脳科学・神経科学が、応用にむけ、企業とどのように関わっていくかというハナシ。
具体的には、NTTデータ経営研究所がこの秋から立ち上げる『応用脳科学コンソーシアム』について、その概要と企業側の期待。
コンソーシアムにおける共同研究が、脳科学の研究成果を、はたしてどれだけ実際の経営につなげられるのかはかなり疑問だが。
企業の側にもっとちゃんとした脳科学に関する知識を提供しようという、コンソーシアムの教育プログラムは、もっともだと思いました。
PL3 Contrasting roles of medial and lateral orbitofrontal cortex in credit assignment and value comparisonこれからの脳科学・神経科学が、応用にむけ、企業とどのように関わっていくかというハナシ。
具体的には、NTTデータ経営研究所がこの秋から立ち上げる『応用脳科学コンソーシアム』について、その概要と企業側の期待。
コンソーシアムにおける共同研究が、脳科学の研究成果を、はたしてどれだけ実際の経営につなげられるのかはかなり疑問だが。
企業の側にもっとちゃんとした脳科学に関する知識を提供しようという、コンソーシアムの教育プログラムは、もっともだと思いました。
Matthew F. S. Rushworth
Rushworthの独壇場。
lateral PFCは意思決定には要りませんかそうですか。
S3-7-2 理論と実験を繋ぐダイナミッククランプlateral PFCは意思決定には要りませんかそうですか。
岡田 真人, 大森 敏明
結構期待していったのですが…
なんというか、わたしには合わなかった(笑)
おひとりめの発表終了時に抜けました。
S3-5-2 個人差の認知神経科学なんというか、わたしには合わなかった(笑)
おひとりめの発表終了時に抜けました。
金井 良太
まったく期待せずにいったのですが…
案の定、わたしには合わなかった(笑)
わたしの嫌いな研究の方向性とはいえ、対象としてる認知機能はすこぶる関係があるので、がまんして聴きにいったのだが。
これだったら、藤井先生のBMI聴きにいったほうが楽しめただろうなぁ。
案の定、わたしには合わなかった(笑)
わたしの嫌いな研究の方向性とはいえ、対象としてる認知機能はすこぶる関係があるので、がまんして聴きにいったのだが。
これだったら、藤井先生のBMI聴きにいったほうが楽しめただろうなぁ。
ポスター
1日目(2010.09.02)
P1-m24 過去の選択が現在の知覚意思決定に与える影響赤石 れい, 坂井 克之
例によってランダム・ドット・モーション課題で、以前の試行での選択と同じ選択をしやすいバイアスについて。
ITIに無関係なRDMとか出して、運動残効の可能性とか排除。
選択にタイムプレッシャーをかけると、バイアスが変化。
過去数試行にさかのぼって影響をみると、5試行前とかからもけっこう影響ある。
逆になぜか、直前の試行からの影響は少ない。(意識的に同じ選択を避けるため?)
あぁ、それから。
去年ポスターで出してた、TMS使ったハナシはどうなりましたかって聞いたんだけど。
「まずはbehaviorできちんと調べる必要があるので、今年はこういう内容」とのことだった。
…ということは…うまくいかなかったのかな(笑)
P1-m25 モデルフリーとモデルベース戦略によるラットの行動選択ITIに無関係なRDMとか出して、運動残効の可能性とか排除。
選択にタイムプレッシャーをかけると、バイアスが変化。
過去数試行にさかのぼって影響をみると、5試行前とかからもけっこう影響ある。
逆になぜか、直前の試行からの影響は少ない。(意識的に同じ選択を避けるため?)
あぁ、それから。
去年ポスターで出してた、TMS使ったハナシはどうなりましたかって聞いたんだけど。
「まずはbehaviorできちんと調べる必要があるので、今年はこういう内容」とのことだった。
…ということは…うまくいかなかったのかな(笑)
船水 章大, 高橋 宏知
左右のノーズポークに対して、異なる強化確率のペアを、全部で4ペア用意。
セッション中に確率ペアを切り替えたときのラットの選択行動が、モデルベースとモデルフリーのどちらの理論で説明できるか。
ここでモデルフリーとは、二選択肢に関する価値関数の比較から行動を決定する、強化学習的モデル。
モデルベースとは、上記のような4種類の確率ペアがあることを知ったうえで、現在がどのスケジュールにあるのかを予測して行動するようなモデル。
尤度でみると、スケジュールの切り替え直後はモデルベースのほうがよくあてはまり、その後だんだんモデルフリーになってくるそうな。
しかし、行動戦略としてはモデルベースのほうが高度なのに、それがだんだん「さっきエサ出たからまたこっち」っていう反応になっていくのは、よく分からん。
単に、スケジュール変更直後のラットのテキトーな選択のばらつきが、たまたまモデルベースのほうでより説明できただけではなかろうか。
シミュレーションによる予測をみると、モデルベースなほうが選択がふらつく傾向があったので。
モデルフリーよりはモデルベースのほうがフィットがよかったからといって、ラットが内的にそういうモデルをもってるかどうかは分からないよね。
相対的な説明率の問題なわけで。
…という指摘をしたのだが、なんとなくはぐらかされてしまった。
セッション中に確率ペアを切り替えたときのラットの選択行動が、モデルベースとモデルフリーのどちらの理論で説明できるか。
ここでモデルフリーとは、二選択肢に関する価値関数の比較から行動を決定する、強化学習的モデル。
モデルベースとは、上記のような4種類の確率ペアがあることを知ったうえで、現在がどのスケジュールにあるのかを予測して行動するようなモデル。
尤度でみると、スケジュールの切り替え直後はモデルベースのほうがよくあてはまり、その後だんだんモデルフリーになってくるそうな。
しかし、行動戦略としてはモデルベースのほうが高度なのに、それがだんだん「さっきエサ出たからまたこっち」っていう反応になっていくのは、よく分からん。
単に、スケジュール変更直後のラットのテキトーな選択のばらつきが、たまたまモデルベースのほうでより説明できただけではなかろうか。
シミュレーションによる予測をみると、モデルベースなほうが選択がふらつく傾向があったので。
モデルフリーよりはモデルベースのほうがフィットがよかったからといって、ラットが内的にそういうモデルをもってるかどうかは分からないよね。
相対的な説明率の問題なわけで。
…という指摘をしたのだが、なんとなくはぐらかされてしまった。
2日目(2010.09.03)
P2-k10 新しい手がかり-標的連合学習時のサル前頭前野ニューロンの反応楠 真琴, John Duncan
対連合学習課題において、新しいペアを入れたときのPFCニューロンの活動について。
逆転学習課題で知られてるみたいに、サルが行動的にペアを学習するにつれて、トライアルベースで発火頻度が変わってくる。
しかし、いつも疑問なんだが、こういう新規学習ってユニット記録でやらなきゃならんもんかね?
同じニューロンをホールドしていられる時間なんて、数時間なんだから。
その活動が行動にどう関連してるかなんて、分からないのでは?
つまり、こういうパラダイムって、宿命的に逸話的な神経活動の報告しかできないように思えるです。
P2-k13 異なる内的認知プロセスを担うセル・アセンブリ・ダイナミクスを解析するための行動課題逆転学習課題で知られてるみたいに、サルが行動的にペアを学習するにつれて、トライアルベースで発火頻度が変わってくる。
しかし、いつも疑問なんだが、こういう新規学習ってユニット記録でやらなきゃならんもんかね?
同じニューロンをホールドしていられる時間なんて、数時間なんだから。
その活動が行動にどう関連してるかなんて、分からないのでは?
つまり、こういうパラダイムって、宿命的に逸話的な神経活動の報告しかできないように思えるです。
中園 智晶, 櫻井 芳雄
セルアセンブリのひとたちのあいだでは、より抽象的な認知機能を発揮中ほど、より大きなアセンブリが活性化されるというのが通説なんだそうな。
そこで、同じ外的刺激によって、異なる抽象度の認知課題をさせるようにラットを訓練しましょう、というハナシ。
課題はすこぶる簡単で、ノーズポークホールが光るんだけど、ある条件ではそれが単純反応課題のインターバルの長さを示す。
別の条件では、それが遅延非見本合わせの位置手がかりになる。
左右の刺激位置の違いから時間という情報に変換する必要のある前者のほうが、より抽象的だろう、ということらしい。
単純系アプローチとしては、ここまでシンプルにしてあることはすばらしいと思う。
(単純すぎて、なんかこういうことってすでにやられてたりしないのかと不安になるけれども。)
ただちょっと気になったのは、アセンブリの大きさをどうやって測るかという問題。
刺入したテトロード間での同期活動を指標にするといってたけど、それは正しい方法なのだろうか。
prefrontalから記録するといっておられたし。
「大きいアセンブリ=物理的に連続した広い領域での同期した活動」ってのは、必ずしも高次連合野にはあてはまらないのでは?
そのへん、アセンブリ屋ではないのでよく分からん。
P2-k14 ラットの系列反応時間課題における順序情報処理*そこで、同じ外的刺激によって、異なる抽象度の認知課題をさせるようにラットを訓練しましょう、というハナシ。
課題はすこぶる簡単で、ノーズポークホールが光るんだけど、ある条件ではそれが単純反応課題のインターバルの長さを示す。
別の条件では、それが遅延非見本合わせの位置手がかりになる。
左右の刺激位置の違いから時間という情報に変換する必要のある前者のほうが、より抽象的だろう、ということらしい。
単純系アプローチとしては、ここまでシンプルにしてあることはすばらしいと思う。
(単純すぎて、なんかこういうことってすでにやられてたりしないのかと不安になるけれども。)
ただちょっと気になったのは、アセンブリの大きさをどうやって測るかという問題。
刺入したテトロード間での同期活動を指標にするといってたけど、それは正しい方法なのだろうか。
prefrontalから記録するといっておられたし。
「大きいアセンブリ=物理的に連続した広い領域での同期した活動」ってのは、必ずしも高次連合野にはあてはまらないのでは?
そのへん、アセンブリ屋ではないのでよく分からん。
石野 誠也, 櫻井 芳雄
P2-k20 行動決定課題および報酬スケジュール課題遂行中のマカクサルの行動成績比較*
瀬戸川 剛, 設楽 宗孝
P2-l02 反応時間課題の訓練による時間推定能力の向上*
角田 吉昭, 筧 慎治
びっくりするほど、タイトルどおり。
ヒト被験者で、時間推定がうまくなりましたよ、という結果。
え、それだけ?
P2-l15 時間弁別課題中のサル線条体の手がかり期の神経細胞の活動特性*ヒト被験者で、時間推定がうまくなりましたよ、という結果。
え、それだけ?
千葉 惇, 稲瀬 正彦
P2-l28 ダイヤモンド電極を用いたサル脳における報酬応答の検出*
吉見 建二, 北澤 茂
ポスター流し読んだだけでは、研究の目的から結果から、さっぱり分からなかった。
なんでダイヤモンド電極なんぞつくったの?
P2-m03 Neural correlates of cognitive dissonance and preference change in the free-choice paradigm*なんでダイヤモンド電極なんぞつくったの?
出馬 圭世, 松元 健二
「あのブドウは酸っぱい」のハナシ。
酸っぱいから選ばなかったのではなく、選ばなかったから酸っぱいと思うのだ、というよくあるストーリー。
P2-m04 Causal interaction between lateral prefrontal cortex and striatum酸っぱいから選ばなかったのではなく、選ばなかったから酸っぱいと思うのだ、というよくあるストーリー。
Xiaochuan Pan, 坂上 雅道
サルにA1-B1-C1とA2-B2-C2という反応規則を覚えさせたあと、新しい刺激を導入したときの神経活動。
単一ニューロン記録についてはもう発表されているので、今回はLFPについてのみ。
結論からいうと、正直いってcausal interactionがどうとかいうハナシではぜんぜんなかった。
まあ予想はしてたけど。
結果はかなりごちゃごちゃで、解釈というか、こじつけは何とでもできる状態。
また、PFCとstriatumの因果関係が、単一ニューロンのときとLFPで逆転していて、ストーリーとしてまとまっていない。
で、困ったからって「結局ループが重要だから、ユニットとLFPで矛盾する結果になるのもしょうがない」とかいわれても、ね。
どうにも納得はできない。
単一ニューロン記録についてはもう発表されているので、今回はLFPについてのみ。
結論からいうと、正直いってcausal interactionがどうとかいうハナシではぜんぜんなかった。
まあ予想はしてたけど。
結果はかなりごちゃごちゃで、解釈というか、こじつけは何とでもできる状態。
また、PFCとstriatumの因果関係が、単一ニューロンのときとLFPで逆転していて、ストーリーとしてまとまっていない。
で、困ったからって「結局ループが重要だから、ユニットとLFPで矛盾する結果になるのもしょうがない」とかいわれても、ね。
どうにも納得はできない。
3日目(2010.09.04)
P3-h03 Temporal relationship between the oscillatory local-filed-potential components and spikes of identified neurons in the rat motor cortex五十嵐 潤, 礒村 宜和, 深井 朋樹
礒村先生が入ってるポスター、いくつかあったのでみたかったが、どれも発表者が居なくて聞けなかった。
で、このポスター読んでたら、ちょうど演者のかたが戻られたので、説明してもらえました。
ハナシとしては、2009のNature Neuroの礒村先生の仕事のデータで、LFPみてみましたというもの。
ただ、結果はなかなか解釈に困るもので。
レバー押し中のM1のLFPにおいて、slow(25-50Hz)およびfast(60-120Hz)なガンマ成分が増加するってとこまではよいのだけど。
その位相が、6層が一番早くて、それが浅い層へと順次移っていくという。
LFPがシナプス後電流の総体だとすると、出力層の位相が一番早いって、ストーリーおかしくね?
また、juxtaのユニット発火のLFP位相へのロックも調べたそうなのだが。
礒村先生の5分類(hold/pre-movement/movement/movement-off/post-movement)でみても、あんまり顕著な位相タイムロックの違いはない。
強いていうと、深い層のニューロンで若干ロックが外れてる?
しかし全体のストーリーとしてはできあがらない。
失礼を承知で、「せっかくのjuxtaなのにその良さがあんまり出てないのでは?」とお聞きすると、たしかにそのとおりとのこと。
interneuronも少ないから、層と細胞形態までちゃんと分かるのに、ニューロン種ごとの違いとかには踏み込めてないし。
もったいない。
はなしをうかがうと、どうもこの研究はこれ以上は進まないかも、と。
最近の微小神経回路業界は、新しい方法論を確立したところで、テーマを模索してる最中というところなのかなぁ…と勝手に納得。
ネズミの頭固定タスクとか、juxtacellularとか、光とか。
P3-h04 第一次視覚野損傷後のサッケード運動機構における網膜視蓋経路の役割で、このポスター読んでたら、ちょうど演者のかたが戻られたので、説明してもらえました。
ハナシとしては、2009のNature Neuroの礒村先生の仕事のデータで、LFPみてみましたというもの。
ただ、結果はなかなか解釈に困るもので。
レバー押し中のM1のLFPにおいて、slow(25-50Hz)およびfast(60-120Hz)なガンマ成分が増加するってとこまではよいのだけど。
その位相が、6層が一番早くて、それが浅い層へと順次移っていくという。
LFPがシナプス後電流の総体だとすると、出力層の位相が一番早いって、ストーリーおかしくね?
また、juxtaのユニット発火のLFP位相へのロックも調べたそうなのだが。
礒村先生の5分類(hold/pre-movement/movement/movement-off/post-movement)でみても、あんまり顕著な位相タイムロックの違いはない。
強いていうと、深い層のニューロンで若干ロックが外れてる?
しかし全体のストーリーとしてはできあがらない。
失礼を承知で、「せっかくのjuxtaなのにその良さがあんまり出てないのでは?」とお聞きすると、たしかにそのとおりとのこと。
interneuronも少ないから、層と細胞形態までちゃんと分かるのに、ニューロン種ごとの違いとかには踏み込めてないし。
もったいない。
はなしをうかがうと、どうもこの研究はこれ以上は進まないかも、と。
最近の微小神経回路業界は、新しい方法論を確立したところで、テーマを模索してる最中というところなのかなぁ…と勝手に納得。
ネズミの頭固定タスクとか、juxtacellularとか、光とか。
加藤 利佳子, 吉田 正俊, 伊佐 正
半側V1を損傷したサルで、損傷とipsiまたはcontraな側の上丘をムシモルで止めたときの視覚誘導性眼球運動成績の検討。
同側上丘にムシモルを入れると、反対視野へのVGSができなくなる。
しかし、その状態で暗室にいれたときの自発性眼球運動をみると、反対視野方向へも眼球は動かせてる。
よって、これは上丘を止めたことによる運動障害ではなく、V1損傷ザルにおける上丘が視知覚に関与してることを示唆する、と。
まあ実験はすごく単純なんだけど、結果はわりとambiguousにみえた。
上丘抑制時に、同側への眼球運動は阻害されないといっても、当然ながらサッケードのカーブは酷くなってるし。
自発性眼球運動も、サッケードベクトルでみるとまんべんなくみえるけど、実際の視点の偏向は、ムシモル前後で変わってる。
どうみても、上丘抑制によって眼球運動に変化が起きている。
よって、観察されたVGS障害を、上丘が担っていた視知覚機能が抑制されたためだと結論付けるには、やや弱いと思った。
P3-h11 一次視覚野損傷後の眼球運動制御における外側頭頂間野の機能的役割同側上丘にムシモルを入れると、反対視野へのVGSができなくなる。
しかし、その状態で暗室にいれたときの自発性眼球運動をみると、反対視野方向へも眼球は動かせてる。
よって、これは上丘を止めたことによる運動障害ではなく、V1損傷ザルにおける上丘が視知覚に関与してることを示唆する、と。
まあ実験はすごく単純なんだけど、結果はわりとambiguousにみえた。
上丘抑制時に、同側への眼球運動は阻害されないといっても、当然ながらサッケードのカーブは酷くなってるし。
自発性眼球運動も、サッケードベクトルでみるとまんべんなくみえるけど、実際の視点の偏向は、ムシモル前後で変わってる。
どうみても、上丘抑制によって眼球運動に変化が起きている。
よって、観察されたVGS障害を、上丘が担っていた視知覚機能が抑制されたためだと結論付けるには、やや弱いと思った。
尾上 浩隆, 吉田 正俊, 伊佐 正
なぜかファーストではなく、吉田先生が説明されていた(笑)
吉田先生いわく、「PETがavailableになったのでやってみた」という研究。
V1破壊ザルに「運動寄り」な課題と「視覚寄り」な課題をブロックデザインでやらせて、関連部位をPETイメージング。
ついでにfunctional connectivity。
まあぶっちゃけて要約すると、頭頂間溝が重要みたいっていうハナシなんだけど…
いかんせんイメージングなので、メカに踏み込んだ考察は少なめ。
吉田先生自身も、
吉田先生いわく、「PETがavailableになったのでやってみた」という研究。
V1破壊ザルに「運動寄り」な課題と「視覚寄り」な課題をブロックデザインでやらせて、関連部位をPETイメージング。
ついでにfunctional connectivity。
まあぶっちゃけて要約すると、頭頂間溝が重要みたいっていうハナシなんだけど…
いかんせんイメージングなので、メカに踏み込んだ考察は少なめ。
吉田先生自身も、
「イメージング手法が面白いのは、いろんな脳領域が光ってきて、謎が謎を呼ぶということ。」
「『じゃあその部位は何をしてんの?』っていう部分は、たとえば電気生理やムシモルつかった抑制で、細胞レベルで調べていくことができる」
とおっしゃっていたので、この結果の意味するところは今後の研究に期待。「『じゃあその部位は何をしてんの?』っていう部分は、たとえば電気生理やムシモルつかった抑制で、細胞レベルで調べていくことができる」
その他
なぜか「ひとりSIRENごっこ」がマイブームだったw感想
なんか、自分の領域に近いハナシに限って、ひどく眠かったような。対象にしてる認知機能のアルゴリズムなんてこれっぽっちも考えてないくせに、
「prefrontalが…をしてて、その情報がparietalにtop-downで伝えられて…」
みたいな勝手な妄想を声高に叫んでいる研究が、最近、どうしてもはなにつくようになってしまって。そういう研究のやりかたって、もう20年前に終焉を迎えてると思うんだけど…
あれやってる方々は、その主張が本当に意味のあるものだと思ってるのでしょうか。
いっぽうで、やはり光や遺伝のハナシは、勢いがあって面白かった。
光遺伝学やjuxtacellularには、本当にこれからの神経科学の未来を感じる。
あたしは専攻する分野を間違えたかも分からんね。
あと、どうでもいいけど、ラストが茂木○一郎になってる『他者から信頼を失ったときに生じる孤独感』っていうポスターがあったんだが。
ポスターを読んでるひとどころか、説明してるひとすらついぞみかけなかったのが、感慨深かった。
悪い意味で。
てか、今回の大会にコイツの名前がはいったポスター2枚でてたんだけどさ。
英語の名前が「Ken Mogi」になってるのは、なんか理由があってのことなの?
研究者としての正式名称がソレなのか?
(※追記:英語名はKen Mogiにしてんだってさ、意味不明ですね。)
悪ふざけだとしたら、神経科学学会は来年から、コイツの発表rejectして良いと思う。
「おたくのようなふざけた名前と髪型のやからには、発表していただけません。」
「てかあんたの研究、神経科学じゃないし。」
ってかんじで。「てかあんたの研究、神経科学じゃないし。」